
Methanotrophy: A Biological Method to Mitigate Global Methane Emission

 , ,
, , 
Abstract
:1. Introduction
2. Sources of Methane Generation
2.1. Emissions from Waste Water
2.2. Emissions from Solid Municipal Waste
2.3. Emissions from Coal Mining and Energy Sectors
2.4. Emissions from Agriculture
2.5. Emissions from Wetlands
2.6. Methane Emission by Methanogens
3. Mitigation Measures of Methane
- ✓
- Installation of vapor recovery units to capture light hydrocarbon vapors that vent out from crude oil storage, followed by conversion of methane to CO2.
- ✓
- Installation of Flash Tank Separators in the production, processing, and transmission units. They can recover methane from tri-ethylene glycol for fuel or sale, thus minimizing venting with water vapor.
- ✓
- Using low-bleed pneumatics or dried air systems eliminates the methane emissions caused by high-bleed pneumatics.
- ✓
- Direct leak detection surveys of facilities to identify and repair leak sources [51].
4. Methanotrophy
4.1. Mechanisms of Methane Oxidation in Methanotrophic Bacteria




4.2. Serine Pathway (Type II Methanotrophs)


4.3. Ribulose Monophosphate Pathway (Type I Methanotrophs)


5. Future Prospects and Applications of Methanotrophs
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, N. A review study on global warming and climate change. Asian J. Adv. Res. 2021, 10, 466–472. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency (USEPA). Overview of Greenhouse Gases. Available online: https://www.epa.gov/ghgemissions/overview-greenhouse-gases (accessed on 3 April 2024).
- University of Washington, United States. Fundamentals of Climate Change. 5 Heat Traps: Earth’s Increasing Greenhouse Effect. Available online: https://uw.pressbooks.pub/fundamentalsofclimatechange/chapter/heat-traps/ (accessed on 3 April 2024).
- U.S. EPA’s: Global Anthropogenic Emissions of Non-CO2 Greenhouse Gases: 1990–2030 Report. Available online: https://nepis.epa.gov/Exe/ZyPURL.cgi?Dockey=P100OHNO.TXT (accessed on 3 April 2024).
- Tseten, T.; Sanjorjo, R.A.; Kwon, M.; Kim, S.W. Strategies to Mitigate Enteric Methane Emissions from Ruminant Animals. J. Microbiol. Biotechnol. 2022, 32, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Ruppel, C.D.; Kessler, J.D. The interaction of climate change and methane hydrates. Rev. Geophys. 2017, 55, 126–168. [Google Scholar] [CrossRef]
- Kennicutt, M.C.; Brooks, J.M.; Bidigare, R.R.; Fay, R.R.; Wade, T.L.; McDonald, T.J. Vent-type taxa in a hydrocarbon seep region on the Louisiana slope. Nature 1985, 317, 351–353. [Google Scholar] [CrossRef]
- de Bruin, G.; ten Veen, J.; Wilpshaar, M.; Versteijlen, N.; Geel, K.; Verweij, H.; Carpentier, S. Origin of shallow gas in the Dutch North Sea—Seismic versus geochemical evidence. Interpretation 2022, 10, SB63–SB76. [Google Scholar] [CrossRef]
- Yurganov, L.; Carroll, D.; Pnyushkov, A.; Polyakov, I.V.; Zhang, H. Ocean stratification and sea-ice cover in Barents and Kara seas modulate sea-air methane flux: Satellite data. Adv. Polar Sci. 2021, 32, 118–140. [Google Scholar]
- Bulavina, A.S.; Vodopianova, V.V.; Zakharenko, V.S.; Makarevich, P.R. Methane migration, sea ice melting and spatial distribution of chlorophyll-a in the Barents Sea. IOP Conf. Ser. Earth Environ. Sci. 2022, 1112, 012019. [Google Scholar] [CrossRef]
- Nisbet, E.G.; Fisher, R.E.; Lowry, D.; France, J.L.; Allen, G.; Bakkaloglu, S.; Broderick, T.J.; Cain, M.; Coleman, M.; Fernandez, J.; et al. Methane Mitigation: Methods to Reduce Emissions, on the Path to the Paris Agreement. Rev. Geophys. 2020, 58, e2019RG000675. [Google Scholar] [CrossRef]
- Sirohi, S.; Michaelowa, A.; Sirohi, S.K. Mitigation options for enteric methane emissions from dairy animals: An Evaluation for potential CDM projects in India. Mitig. Adapt. Strateg. Glob. Change 2007, 12, 259–274. [Google Scholar] [CrossRef]
- Koo, C.W.; Rosenzweig Amy, C. Biochemistry of aerobic biological methane oxidation. Chem. Soc. Rev. 2021, 50, 3424. [Google Scholar] [CrossRef]
- Scheehle, E.A.; Kruger, D. Global anthropogenic methane and nitrous oxide emissions. Energy J. 2006, 27 (Suppl. S3), 33–44. [Google Scholar] [CrossRef]
- Höglund-Isaksson, L. Global anthropogenic methane emissions 2005–2030: Technical mitigation potentials and costs. Atmos. Chem. Phys. 2012, 12, 9079–9096. [Google Scholar] [CrossRef]
- Kirschke, S.; Bousquet, P.; Ciais, P.; Saunois, M.; Canadell, J.G.; Dlugokencky, E.J.; Bergamaschi, P.; Bergmann, D.; Blake, D.R.; Bruhwiler, L.; et al. Three decades of global methane sources and sinks. Nat. Geosci. 2013, 6, 813–823. [Google Scholar] [CrossRef]
- Reay, D.S.; Smith, K.; Hewitt, C. Methane: Importance, sources and sinks. In Greenhouse Gas Sinks; CAB International: Wallingford, UK, 2007; Volume 21, pp. 143–151. [Google Scholar]
- Meinshausen, M.; Meinshausen, N.; Hare, W.; Raper, S.C.; Frieler, K.; Knutti, R.; Frame, D.J.; Allen, M.R. Greenhouse-gas emission targets for limiting global warming to 2 C. Nature 2009, 458, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- GMAO Chemical Forecasts and GEOS–CHEM NRT Simulations for ICARTT (top) and Randy Kawa, NASA GSFC Atmospheric Chemistry and Dynamics Branch (Lower). Available online: https://web.archive.org/web/20050313053747/http://earthobservatory.nasa.gov/Newsroom/NewImages/images.php3?img_id=16827 (accessed on 3 April 2024).
- Nisbet, E.G.; Manning, M.R.; Dlugokencky, E.J.; Fisher, R.E.; Lowry, D.; Michel, S.E.; Myhre, C.L.; Platt, S.M.; Allen, G.; Bousquet, P.; et al. Very strong atmospheric methane growth in the 4 years 2014–2017: Implications for the Paris Agreement. Glob. Biogeochem. Cycles 2019, 33, 318–342. [Google Scholar] [CrossRef]
- Stein, T.; Spinrad, R. Increase in atmospheric methane set another record during 2021. NOAA Res. News. 2022, 497–6288. [Google Scholar]
- Tubiello, F.N.; Conchedda, G.; Wanner, N.; Federici, S.; Rossi, S.; Grassi, G. Carbon emissions and removals from forests: New estimates, 1990–2020. Earth Syst. Sci. Data 2021, 13, 1681–1691. [Google Scholar] [CrossRef]
- Global Monitoring Laboratory. Trends in CO2, CH4, N2O, SF6. Available online: https://gml.noaa.gov/ccgg/trends_ch4/ (accessed on 3 April 2024).
- The Methane Thread. Available online: https://peakoil.com/forums/viewtopic.php?p=1492258 (accessed on 3 April 2024).
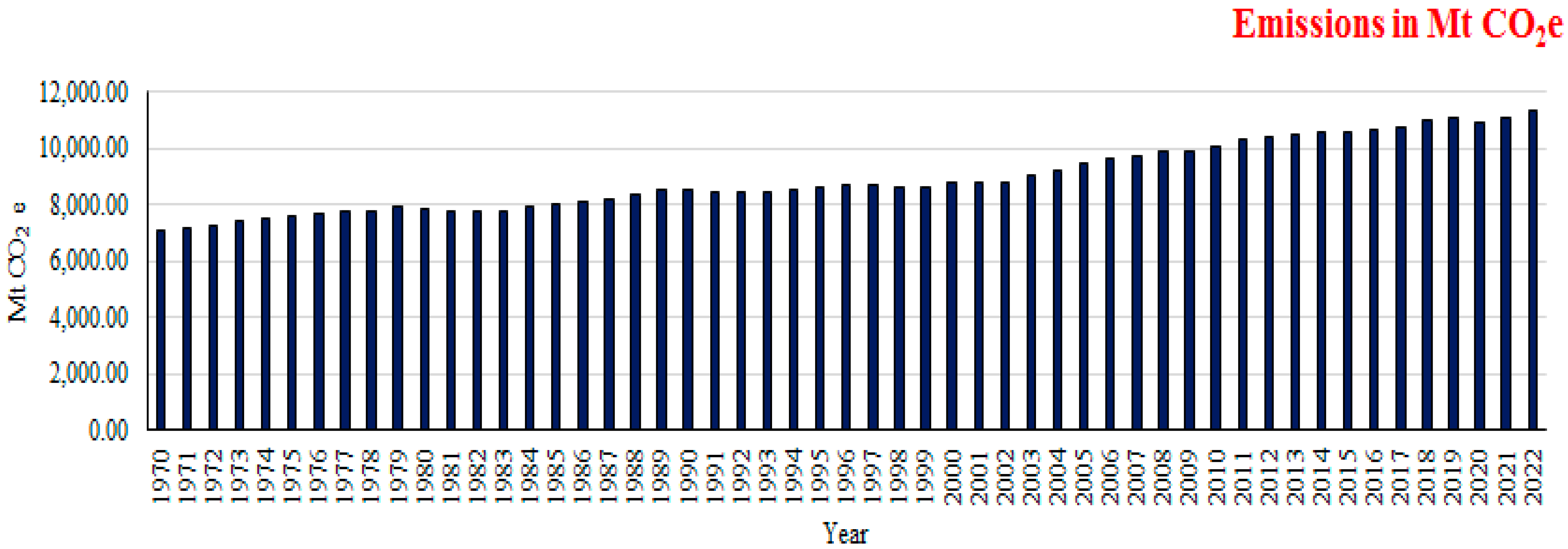
- Annual Methane Emissions Worldwide 1970–2022. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/1298441/annual-global-methane-emissions/ (accessed on 3 February 2024).
- Parry, I.W.; Black, M.S.; Minnett, D.N.; Mylonas, M.V.; Vernon, N. How to Cut Methane Emissions; International Monetary Fund: Bretton Woods, NH, USA, 2022. [Google Scholar]
- Global Methane Initiative (GMI). Global Methane Emissions and Mitigation Opportunities. 2011. Available online: https://www.globalmethane.org/documents/analysis_fs_en.pdf (accessed on 5 April 2024).
- Global Methane Tracker 2022. Available online: https://www.iea.org/reports/global-methane-tracker-2022/overview (accessed on 3 February 2024).
- Hopkins, F.M.; Ehleringer, J.R.; Bush, S.E.; Duren, R.M.; Miller, C.E.; Lai, C.T.; Hsu, Y.K.; Carranza, V.; Randerson, J.T. Mitigation of methane emissions in cities: How new measurements & partnerships can contribute to emission reduction strategies. Earth’s Future 2016, 4, 408–425. [Google Scholar]
- Kulkarni, B.N.; Kulkarni, N.B.; Anantharama, V. An empirical analysis of surface-level methane emission from anthropogenic sources in India. J. Clean. Prod. 2022, 346, 131101. [Google Scholar] [CrossRef]
- NASA. Methane | Vital Signs—Climate Change. Available online: https://climate.nasa.gov/vital-signs/methane/ (accessed on 5 April 2024).
- El-Fadel, M.; Massoud, M. Methane emissions from wastewater management. Environ. Pollut. 2001, 114, 177–185. [Google Scholar] [CrossRef]
- Foley, J.; Yuan, Z.; Lant, P. Dissolved methane in rising main sewer systems: Field measurements and simple model development for estimating greenhouse gas emissions. Water Sci. Technol. 2009, 60, 2963–2971. [Google Scholar] [CrossRef]
- Guisasola, A.; de Haas, D.; Keller, J.; Yuan, Z. Methane formation in sewer systems. Water Res. 2008, 42, 1421–1430. [Google Scholar] [CrossRef]
- Daelman, M.R.; van Voorthuizen, E.M.; van Dongen, U.G.; Volcke, E.I.; van Loosdrecht, M.C. Methane emission during municipal wastewater treatment. Water Res. 2012, 46, 3657–3670. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Q.; Li, J.; Wang, Z. Methane in wastewater treatment plants: Status, characteristics, and bioconversion feasibility by methane oxidizing bacteria for high value-added chemicals production and wastewater treatment. Water Res. 2021, 198, 117122. [Google Scholar] [CrossRef]
- Lee, O.K.; Hur, D.H.; Nguyen, D.T.; Lee, E.Y. Metabolic engineering of methanotrophs and its application to production of chemicals and biofuels from methane. Biofuels Bioprod. Biorefining 2016, 10, 848–863. [Google Scholar] [CrossRef]
- Nugroho, W. Application of CO2 Microbubble to Enhance Methane Production in Anaerobic Digestion of Food Waste. Doctoral Dissertation, University of Sheffield, Sheffield, UK, 2021. [Google Scholar]
- Wang, X.; Lei, Z.; Zhang, Z.; Shimizu, K.; Lee, D.J.; Khanal, S.K. Use of nanobubble water bioaugmented anaerobically digested sludge for high-efficacy energy production from high-solids anaerobic digestion of corn straw. Sci. Total Environ. 2023, 863, 160825. [Google Scholar] [CrossRef]
- Agarwal, A.; Ng, W.J.; Liu, Y. Principle and applications of microbubble and nanobubble technology for water treatment. Chemosphere 2011, 84, 1175–1180. [Google Scholar] [CrossRef]
- Moldal, D. Wastewater Treatment, Powered by Biogas. 2021. Available online: https://www.waterworld.com/wastewater/treatment/article/14214366/2112wwcas (accessed on 5 April 2024).
- Foley, J.; Lant, P.A. Direct Methane and Nitrous Oxide Emissions from Full-Scale Wastewater Treatment Systems; The University of Queensland: Brisbane, Australia, 2009. [Google Scholar]
- Campos Gómez, J.L.; Valenzuela-Heredia, D.; Pedrouso Fuentes, A.; Val del Río, M.Á.; Belmonte, M.; Mosquera Corral, A. Greenhouse gases emissions from wastewater treatment plants: Minimization, treatment, and prevention. J. Chem. 2016, 2016, 3796352. [Google Scholar] [CrossRef]
- Singh, C.K.; Kumar, A.; Roy, S.S. Quantitative analysis of the methane gas emissions from municipal solid waste in India. Sci. Rep. 2018, 8, 2913. [Google Scholar] [CrossRef]
- Cai, B.-F.; Liu, J.-G.; Gao, Q.-X.; Nie, X.-Q.; Cao, D.; Liu, L.-C.; Zhou, Y.; Zhang, Z.-S. Estimation of methane emissions from municipal solid waste landfills in China based on point emission sources. Adv. Clim. Change Res. 2014, 5, 81–91. [Google Scholar]
- Lando, A.T.; Rahim, I.R.; Sari, K.; Djamaluddin, I.; Arifin, A.N.; Sari, A.M. Estimation of methane emissions from municipal solid waste landfill in makassar city based on ipcc waste model. IOP Conf. Ser. Earth Environ. Sci. 2021, 841, 012002. [Google Scholar] [CrossRef]
- Managing Methane in the Waste Sector, A Growing Problem, RMI. Available online: https://rmi.org/our-work/climate-aligned-industries/managing-methane-in-the-waste-sector/ (accessed on 6 April 2024).
- Methane Emissions from Open Dumpsites in India, 21 September 2023. Available online: https://www.cseindia.org/methane-emissions-from-open-dumpsites-in-india-11864 (accessed on 6 April 2024).
- Yusuf, R.O.; Noor, Z.Z.; Abba, A.H.; Hassan, M.A.A.; Din, M.F.M. Methane emission by sectors: A comprehensive review of emission sources and mitigation methods. Renew. Sustain. Energy Rev. 2012, 16, 5059–5070. [Google Scholar] [CrossRef]
- Pandey, B.; Gautam, M.; Agrawal, M. Greenhouse gas emissions from coal mining activities and their possible mitigation strategies. In Environmental Carbon Footprints; Butterworth-Heinemann: Oxford, UK, 2018; pp. 259–294. [Google Scholar]
- Karakurt, I.; Aydin, G.; Aydiner, K. Sources and mitigation of methane emissions by sectors: A critical review. Renew. Energy 2012, 39, 40–48. [Google Scholar] [CrossRef]
- Merrill, T.W.; Schizer, D.M. The shale oil and gas revolution, hydraulic fracturing, and water contamination: A regulatory strategy. Minn. L. Rev. 2013, 98, 145. [Google Scholar] [CrossRef]
- Methane Abatement, Global Methane Emissions from the Energy Sector, 2000–2022. Available online: https://www.iea.org/energy-system/fossil-fuels/methane-abatement (accessed on 6 April 2024).
- U.S. Environmental Protection Agency (USEPA). Why is EPA Concerned about CMM? Available online: https://www.epa.gov/cmop/about-coal-mine-methane (accessed on 6 April 2024).
- Xu, Z.; Zheng, Y.; Wu, Y. Paving the way for sustainable agriculture: An analysis of evolution and driving forces of methane emissions reduction in China. Resour. Conserv. Recycl. 2024, 202, 107392. [Google Scholar] [CrossRef]
- Hegde, U.; Chang, T.C.; Yang, S.S. Methane and carbon dioxide emissions from Shan-Chu-Ku landfill site in northern Taiwan. Chemosphere 2003, 52, 127585. [Google Scholar] [CrossRef]
- Howarth, R.W. Methane and Climate Change. In Environmental Impacts from Development of Unconventional Oil and Gas Reserves; John, F., Stolz, W., Griffin, M., Bain, D.J., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Islam, M.; Lee, S.S. Advanced estimation and mitigation strategies: A cumulative approach to enteric methane abatement from ruminants. J. Anim. Sci. Technol. 2019, 61, 122–137. [Google Scholar] [CrossRef]
- Sejian, V.; Naqvi, S.M. Livestock and climate change: Mitigation strategies to reduce methane production. Greenh. Gases-Capturing Util. Reduct. 2012, 9, 255–276. [Google Scholar]
- Sejian, V.; Lal, R.; Lakritz, J.; Ezeji, T. Measurement and Prediction of Enteric Methane Emission. Int. J. Biometeorol. 2011, 55, 1–16. [Google Scholar] [CrossRef]
- Malerba, M.E.; de Kluyver, T.; Wright, N.; Schuster, L.; Macreadie, P.I. Methane emissions from agricultural ponds are underestimated in national greenhouse gas inventories. Commun. Earth Environ. 2022, 3, 306. [Google Scholar] [CrossRef]
- Smith, P.; Reay, D.; Smith, J. Agricultural methane emissions and the potential formitigation. Philos. Trans. R. Soc. A 2021, 379, 20200451. [Google Scholar] [CrossRef] [PubMed]
- Basso, L.S.; Marani, L.; Gatti, L.V.; Miller, J.B.; Gloor, M.; Melack, J.; Cassol, H.L.; Tejada, G.; Domingues, L.G.; Arai, E.; et al. Amazon methane budget derived from multi-year airborne observations highlights regional variations in emissions. Commun. Earth Environ. 2021, 2, 246. [Google Scholar] [CrossRef]
- Saunois, M.; Stavert, A.R.; Poulter, B.; Bousquet, P.; Canadell, J.G.; Jackson, R.B.; Raymond, P.A.; Dlugokencky, E.J.; Houweling, S.; Patra, P.K.; et al. The global methane budget 2000–2017. Earth Syst. Sci. Data Discuss. 2019, 2019, 1–36. [Google Scholar] [CrossRef]
- Zhang, B.; Tian, H.; Lu, C.; Chen, G.; Pan, S.; Anderson, C.; Poulter, B. Methane emissions from global wetlands: An assessment of the uncertainty associated with various wetland extent data sets. Atmos. Environ. 2017, 165, 310–321. [Google Scholar] [CrossRef]
- Basu, A. An Analysis of the Global Atmospheric Methane Budget under Different Climates; Forschungszentrum Jülich: Jülich, Germany, 2013. [Google Scholar]
- Peng, S.; Lin, X.; Thompson, R.L.; Xi, Y.; Liu, G.; Hauglustaine, D.; Lan, X.; Poulter, B.; Ramonet, M.; Saunois, M.; et al. Wetland emission and atmospheric sink changes explain methane growth in 2020. Nature 2022, 612, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Murguia-Flores, F.; Jaramillo, V.J.; Gallego-Sala, A. Assessing methane emissions from tropical wetlands: Uncertainties from natural variability and drivers at the global scale. Glob. Biogeochem. Cycles 2023, 37, e2022GB007601. [Google Scholar] [CrossRef]
- Ortiz-Llorente, M.J.; Alvarez-Cobelas, M. Comparison of biogenic methane emissions from unmanaged estuaries, lakes, oceans, rivers and wetlands. Atmos. Environ. 2012, 59, 328–337. [Google Scholar] [CrossRef]
- Rosentreter, J.A.; Borges, A.V.; Deemer, B.R.; Holgerson, M.A.; Liu, S.; Song, C.; Melack, J.; Raymond, P.A.; Duarte, C.M.; Allen, G.H.; et al. Half of global methane emissions come from highly variable aquatic ecosystem sources. Nat. Geosci. 2021, 14, 225–230. [Google Scholar] [CrossRef]
- GMI Policymaker Framework for Addressing Methane Emissions. Available online: www.globalmethane.org (accessed on 12 February 2024).
- Conrad, R. Microbial ecology of methanogens and methanotrophs. Adv. Agron. 2007, 96, 1–63. [Google Scholar]
- Chai, X.; Tonjes, D.J.; Mahajan, D. Methane emissions as energy reservoir: Context, scope, causes and mitigation strategies. Prog. Energy Combust. Sci. 2016, 56, 33–70. [Google Scholar] [CrossRef]
- Dutta, H.; Dutta, A. The microbial aspect of climate change. Energy Ecol. Environ. 2016, 1, 209–232. [Google Scholar] [CrossRef]
- Hedderich, R.; Whitman, W.B. Physiology and biochemistry of the methane-producing Archaea. Prokaryotes 2006, 2, 1050–1079. [Google Scholar]
- Hook, S.E.; Wright, A.D.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef] [PubMed]
- Deppenmeier, U.; Müller, V.; Gottschalk, G. Pathways of energy conservation in methanogenicarchaea. Arch. Microbiol. 1996, 165, 149–163. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, S.Y.; Kim, P.J.; Madsen, E.L.; Jeon, C.O. Methane emission and dynamics of methanotrophic and methanogenic communities in a flooded rice field ecosystem. FEMS Microbiol. Ecol. 2014, 88, 195–212. [Google Scholar] [CrossRef]
- Angle, J.C.; Morin, T.H.; Solden, L.M.; Narrowe, A.B.; Smith, G.J.; Borton, M.A.; Rey-Sanchez, C.; Daly, R.A.; Mirfenderesgi, G.; Hoyt, D.W.; et al. Methanogenesis in oxygenated soils is a substantial fraction of wetland methane emissions. Nat. Commun. 2017, 8, 1567. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Chen, Y.; Murrell, J.C.; Jiang, P.; Zhang, C.; Xing, X.H.; Smith, T.J. Methanotrophs: Multifunctional Bacteria with Promising Applications in Environmental Bioengineering. In Comprehensive Biotechnology, 2nd ed.; Murray, M.-Y., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 249–262. ISBN 9780080885049. [Google Scholar]
- Murray, R.M.; Bryant, A.M.; Leng, R.A. Rates of production of methane in the rumen and large intestine of sheep. Br. J. Nutr. 1976, 36, 1–4. [Google Scholar] [CrossRef]
- Morkhade, S.J.; Bansod, A.P.; Kolaskar, A.G.; Thakare, S.O. A complete review on: Methanogens methane producers of rumen and abatement strategies-biotechnology and microbiological strategies review. Int. J. Vet. Sci. Anim. Husb. 2020, 5, 11–17. [Google Scholar]
- Sawyer, W.J.; Genina, I.; Brenneis, R.J.; Feng, H.; Li, Y.; Luo, S.X. Methane emissions and global warming: Mitigation technologies, policy ambitions, and global efforts. MIT Sci. Policy Rev. 2022, 3, 73–84. [Google Scholar] [CrossRef]
- Króliczewska, B.; Pecka-Kiełb, E.; Bujok, J. Strategies used to reduce methane emissions from ruminants: Controversies and issues. Agriculture 2023, 13, 602. [Google Scholar] [CrossRef]
- Mirzaei-Aghsaghali, A.; Maheri-Sis, N. Factors affecting mitigation of methane emission from ruminants I: Feeding strategies. Asian J. Anim. Vet. Adv. 2011, 6, 888–908. [Google Scholar] [CrossRef]
- Palangi, V.; Taghizadeh, A.; Abachi, S.; Lackner, M. Strategies to mitigate enteric methane emissions in ruminants: A review. Sustainability 2022, 14, 13229. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Abdalla, A.L.; Alvarez, C.; Arndt, C.; Becquet, P.; Benchaar, C.; Berndt, A.; Mauricio, R.M.; McAllister, T.A.; et al. Invited review: Current enteric methane mitigation options. J. Dairy Sci. 2022, 105, 9297–9326. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.J.; Hristov, A.N.; Henderson, B.; Makkar, H.; Oh, J.; Lee, C.; Meinen, R.; Montes, F.; Ott, T.; Firkins, J.; et al. Technical options for the mitigation of direct methane and nitrous oxide emissions from livestock: A review. Animal 2013, 7 (Suppl. S2), 220–234. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.; Kebreab, E. Recent advances in feed additives with the potential to mitigate enteric methane emissions from ruminant livestock. J. Soil Water Conserv. 2023, 78, 111–123. [Google Scholar] [CrossRef]
- Mu, L.; Wang, Y.; Xu, F.; Li, J.; Tao, J.; Sun, Y.; Song, Y.; Duan, Z.; Li, S.; Chen, G. Emerging Strategies for Enhancing Propionate Conversion in Anaerobic Digestion: A Review. Molecules 2023, 28, 3883. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.B.; Okada, D.Y.; Delforno, T.P.; Foresti, E. Methane-oxidizing archaea, aerobic methanotrophs and nitrifiers coexist with methane as the sole carbon source. Int. Biodeterior. Biodegrad. 2019, 138, 57–62. [Google Scholar] [CrossRef]
- Abdelbagi, M.; Ridwan, R.; Fidriyanto, R.; Jayanegara, A. Effects of probiotics and encapsulated probiotics on enteric methane emission and nutrient digestibility in vitro. IOP Conf. Ser. Earth Environ. Sci. 2021, 788, 012050. [Google Scholar] [CrossRef]
- Nguyen, S.H.; Nguyen, H.D.; Hegarty, R.S. Defaunation and its impacts on ruminal fermentation, enteric methane production and animal productivity. Livest. Res. Rural. Dev. 2020, 32, 1–9. [Google Scholar]
- Yáñez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 162946. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.; et al. An inhibitor persistently decreased enteric methane emission from dairy cows with no negative effect on milk production. Proc. Natl. Acad. Sci. USA 2015, 112, 10663–10668. [Google Scholar] [CrossRef] [PubMed]
- Neue, H.U. Methane emission from rice fields. Bioscience 1993, 43, 466–474. [Google Scholar] [CrossRef]
- Sass, R.L.; Fisher, F.M.; Wang, Y.B.; Turner, F.T.; Jund, M.F. Methane emission from rice fields: The effect of floodwater management. Glob. Biogeochem. Cycles 1992, 6, 249–262. [Google Scholar] [CrossRef]
- Silva, A.J.; Coelho, E.F. Estimation of water percolation by different methods using TDR. Rev. Bras. De Ciência Do Solo 2014, 38, 73–81. [Google Scholar] [CrossRef]
- Ward, N.D.; Bianchi, T.S.; Martin, J.B.; Quintero, C.J.; Sawakuchi, H.O.; Cohen, M.J. Pathways for methane emissions and oxidation that influence the net carbon balance of a subtropical cypress swamp. Front. Earth Sci. 2020, 8, 573357. [Google Scholar] [CrossRef]
- Samanta, D.; Sani, R.K. Methane oxidation via chemical and biological methods: Challenges and solutions. Methane 2023, 2, 279–303. [Google Scholar] [CrossRef]
- Sainju, U.M.; Ghimire, R.; Dangi, S. Soil carbon dioxide and methane emissions and carbon balance with crop rotation and nitrogen fertilization. Sci. Total Environ. 2021, 775, 145902. [Google Scholar] [CrossRef]
- Hilgert, J.E.; Amon, B.; Amon, T.; Belik, V.; Dragoni, F.; Ammon, C.; Cárdenas, A.; Petersen, S.O.; Herrmann, C. Methane emissions from livestock slurry: Effects of storage temperature and changes in chemical composition. Sustainability 2022, 14, 9934. [Google Scholar] [CrossRef]
- Banks, J.; Force, C.A. Barriers and Opportunities for Reducing Methane Emmissions from Coal Mines. In Clean Air Task Force; US Environmental Protection Agency and the Global Methane Initiative; Raven Ridge Resources, 2012; pp. 1–22. Available online: http://www.ravenridge.com/ (accessed on 13 March 2024).
- Kajaste-Rudnitskaja, R.; Hurme, M.; Oinas, P. Methanol-Managing greenhouse gas emissions in the production chain by optimizing the resource base. AIMS Energy 2018, 6, 1074–1102. [Google Scholar] [CrossRef]
- Mines, U.C. Identifying Opportunities for Methane Recovery at US Coal Mines: Profiles of Selected Gassy. Available online: https://www.epa.gov/sites/default/files/2016-03/documents/profiles_2008_final.pdf (accessed on 7 April 2024).
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef] [PubMed]
- Whittenbury, R.; Dalton, H. The Methylotrophic Bacteria. In The Prokaryotes; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1981. [Google Scholar]
- Gal’chenko, V.F.; Andreev, L.V. Taxonomy of obligate methylo-trophs. In Microbial Growth on C1 Compounds; Crawford, R.L., Hanson, R.S., Eds.; American Society for Microbiology: Washington, DC, USA, 1984; pp. 269–281. [Google Scholar]
- Hanson, R.S. Introduction. In Methane and Methanol Utilizers; Murrell, J.C., Dalton, H., Eds.; Plenum Press: New York, NY, USA, 1992; pp. 1–22. [Google Scholar]
- McDonald, I.R.; Bodrossy, L.; Chen, Y.; Murrell, J.C. Molecular ecology techniques for the study of aerobic methanotrophs. Appl Environ. Microbiol. 2008, 74, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Murrell, J.C. Methanotrophy/methane oxidation. Encycl. Microbiol. 2009, 3, 293–298. [Google Scholar]
- Li, J.; Liu, C.; Wu, N.; He, X.; Hao, X.; Li, F.; Chen, Y.; Meng, Q. The Variation of Microbial (Methanotroph) Communities in Marine Sediments Due to Aerobic Oxidation of Hydrocarbons. J. Ocean. Univ. China 2021, 20, 553–561. [Google Scholar] [CrossRef]
- Guerrero-Cruz, S.; Vaksmaa, A.; Horn, M.A.; Niemann, H.; Pijuan, M.; Ho, A. Methanotrophs: Discoveries, Environmental Relevance, and a Perspective on Current and Future Applications. Front. Microbiol. 2021, 12, 678057. [Google Scholar] [CrossRef] [PubMed]
- Hogendoorn, C.; Pol, A.; Nuijten, G.H.L.; Op den Camp, H.J.M. Methanol production by “Methylacidiphilum fumariolicum” solv under different growth conditions. Appl. Environ. Microbiol. 2020, 86, e01188-20. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.; Hernández, M.; Gessesse, A.; Murrell, J.C.; Øvreås, L. A novel moderately thermophilic facultative methylotroph within the class Alphaproteobacteria. Microorganisms 2021, 9, 903077. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M.G.; Kumaresan, D.; Heimann, K.; Caetano, N.S.; Visvanathan, C.; Parthiba Karthikeyan, O. Editorial: Methane: A bioresource for fuel and biomolecules. Front. Environ. Sci. 2020, 8, 9. [Google Scholar] [CrossRef]
- Ross, M.O.; Rosenzweig, A.C. A tale of two methane monooxygenases. J. Biol. Inorg. Chem. 2017, 22, 307–319. [Google Scholar] [CrossRef]
- Lieven, C.; Petersen, L.A.H.; Jørgensen, S.B.; Gernaey, K.V.; Herrgard, M.J.; Sonnenschein, N. A Genome-Scale Metabolic Model for Methylococcus capsulatus (Bath) Suggests Reduced Efficiency Electron Transfer to the Particulate Methane Monooxygenase. Front. Microbiol. 2018, 9, 2947. [Google Scholar] [CrossRef]
- Anthony, C. Bacterial oxidation of methane and methanol. Adv. Microb. Physiol. 1986, 27, 113–210. [Google Scholar] [PubMed]
- Kinnaman, F.S.; Valentine, D.L.; Tyler, S.C. Carbon and hydrogen isotope fractionation associated with the aerobic microbial oxidation of methane, ethane, propane and butane. Geochim. Cosmochim. Acta 2007, 71, 271–283. [Google Scholar] [CrossRef]
- Semrau, J.D. Bioremediation via Methanotrophy: Overview of Recent Findings and Suggestions for Future Research. Front. Microbiol. 2011, 2, 209. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.P.; Dickinson, D.; Chase, H.A. Methanotrophs, Methylosinus trichosporium OB3b, sMMO, and their application to bioremediation. Crit. Rev. Microbiol. 1998, 24, 335–373. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Involved Characteristics | Gammmaproteobacteria (Type 1 Methanotrophs) | Alphaproteobacteria (Type 2 Methanotrophs) |
|---|---|---|
| Arrangement within cell membrane: | Disc-shaped utricles in bundle forms | Coupled membrane along the periphery |
| Assimilation process: (Carbon compounds) | Undergo Ribulose monophosphate pathway | Undergo serine pathway |
| Type of organism include: | Paraphyletic | Oligotrophic |
| Species under category: | Methylococcus capsulatus, Methylobacter marinus, Methylomonas methanica | Methylosinustrichosporium, Methlocystisminimus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rani, A.; Pundir, A.; Verma, M.; Joshi, S.; Verma, G.; Andjelković, S.; Babić, S.; Milenković, J.; Mitra, D. Methanotrophy: A Biological Method to Mitigate Global Methane Emission. Microbiol. Res. 2024, 15, 634-654. https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres15020042
Rani A, Pundir A, Verma M, Joshi S, Verma G, Andjelković S, Babić S, Milenković J, Mitra D. Methanotrophy: A Biological Method to Mitigate Global Methane Emission. Microbiology Research. 2024; 15(2):634-654. https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres15020042
Chicago/Turabian StyleRani, Anju, Aarushi Pundir, Medhashree Verma, Samiksha Joshi, Geeta Verma, Snežana Andjelković, Snežana Babić, Jasmina Milenković, and Debasis Mitra. 2024. "Methanotrophy: A Biological Method to Mitigate Global Methane Emission" Microbiology Research 15, no. 2: 634-654. https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres15020042