
Identification of HLA-A*11:01 and A*02:01-Restricted EBV Peptides Using HLA Peptidomics

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Plasmids
2.3. Generation of HLA-A*11:01/HLA-A*02:01-Transfected NPC Cell Lines
2.4. Immunopeptidome Sample Preparation and Collection
2.5. MHC-Peptide Immunoprecipitation
2.6. Western Blot
2.7. LC-MS/MS and Data Analysis
2.8. Peptide Synthesis
2.9. Human Peripheral Blood Mononuclear Cell (PBMC) Isolation and HLA Typing
2.10. Peptide-Specific T Cell Line Generation
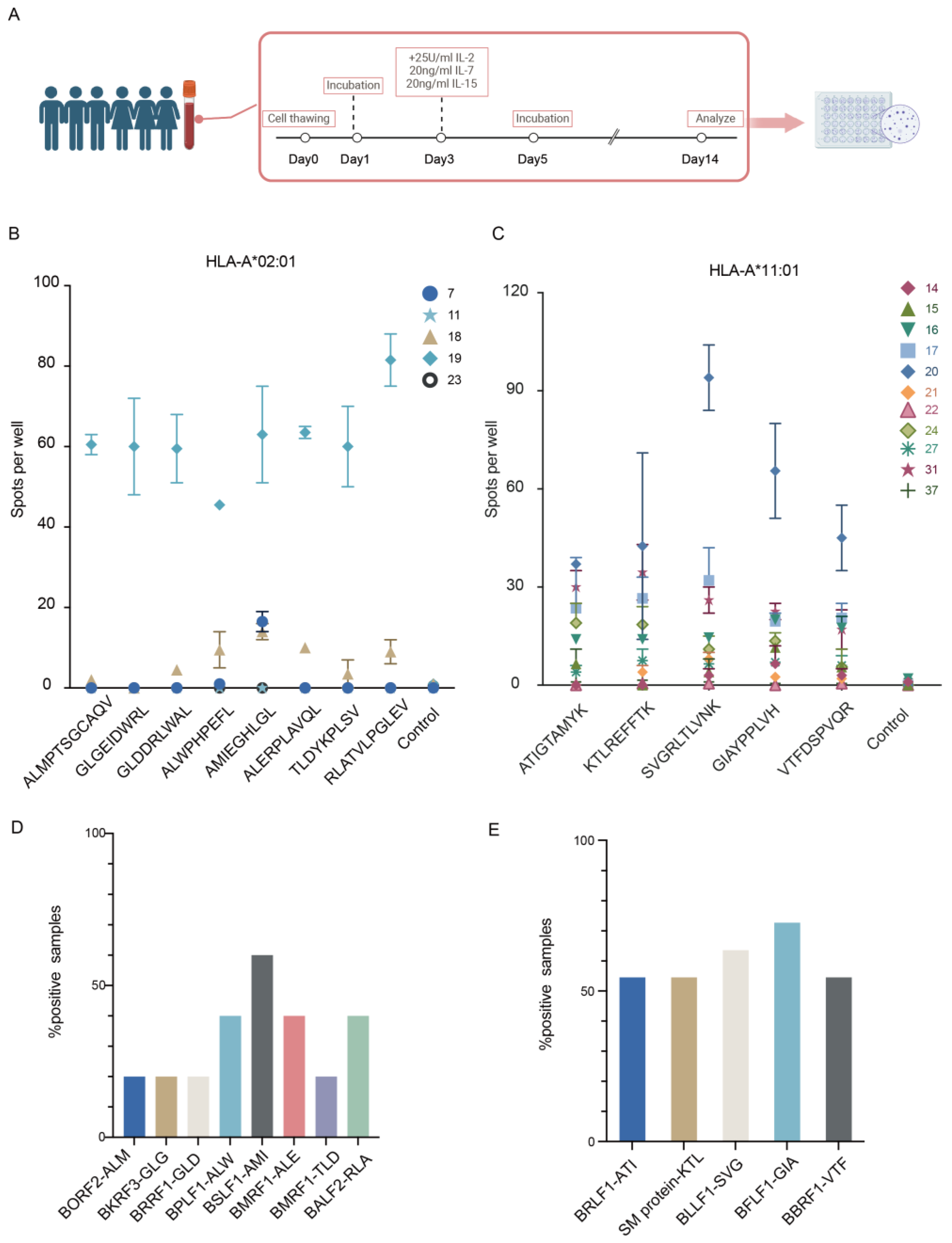
2.11. IFN-γ ELISPOT Assays
2.12. Flow Cytometry
3. Results
3.1. Subsection
3.1.1. Construction of Cell Line with Tagged HLA
3.1.2. Isolation and Identification of EBV Peptides Presented on NPC Cell Lines
3.1.3. Detection of High Confident EBV Epitopes
3.1.4. Immunogenicity of Identified EBV Peptides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein–Barr Virus: More than 50 Years Old and Still Providing Surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Taylor, G.S.; Long, H.M.; Brooks, J.M.; Rickinson, A.B.; Hislop, A.D. The Immunology of Epstein-Barr Virus–Induced Disease. Annu. Rev. Immunol. 2015, 33, 787–821. [Google Scholar] [CrossRef]
- Yin, H.; Qu, J.; Peng, Q.; Gan, R. Molecular Mechanisms of EBV-Driven Cell Cycle Progression and Oncogenesis. Med. Microbiol. Immunol. 2019, 208, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Gullo, C.; Low, W.K.; Teoh, G. Association of Epstein-Barr Virus with Nasopharyngeal Carcinoma and Current Status of Development of Cancer-Derived Cell Lines. Ann. Acad. Med. Singap. 2008, 37, 769–777. [Google Scholar] [CrossRef]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein–Barr Virus Infection and Nasopharyngeal Carcinoma. Philos. Trans. R. Soc. B 2017, 372, 20160270. [Google Scholar] [CrossRef] [PubMed]
- Arvey, A.; Tempera, I.; Tsai, K.; Chen, H.-S.; Tikhmyanova, N.; Klichinsky, M.; Leslie, C.; Lieberman, P.M. An Atlas of the Epstein-Barr Virus Transcriptome and Epigenome Reveals Host-Virus Regulatory Interactions. Cell Host Microbe 2012, 12, 233–245. [Google Scholar] [CrossRef]
- Su, Z.Y.; Siak, P.Y.; Leong, C.-O.; Cheah, S.-C. The Role of Epstein–Barr Virus in Nasopharyngeal Carcinoma. Front. Microbiol. 2023, 14, 1116143. [Google Scholar] [CrossRef]
- Yap, L.F.; Wong, A.K.C.; Paterson, I.C.; Young, L.S. Functional Implications of Epstein-Barr Virus Lytic Genes in Carcinogenesis. Cancers 2022, 14, 5780. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhu, N.; Zheng, J.; Peng, Y.; Zeng, M.-S.; Deng, K.; Duan, C.; Yuan, Y. EBV Abortive Lytic Cycle Promotes Nasopharyngeal Carcinoma Progression through Recruiting Monocytes and Regulating Their Directed Differentiation. PLoS Pathog. 2024, 20, e1011934. [Google Scholar] [CrossRef]
- Hu, L.; Lin, Z.; Wu, Y.; Dong, J.; Zhao, B.; Cheng, Y.; Huang, P.; Xu, L.; Xia, T.; Xiong, D.; et al. Comprehensive Profiling of EBV Gene Expression in Nasopharyngeal Carcinoma through Paired-End Transcriptome Sequencing. Front. Med. 2016, 10, 61–75. [Google Scholar] [CrossRef]
- Ramayanti, O.; Juwana, H.; Verkuijlen, S.A.M.W.; Adham, M.; Pegtel, M.D.; Greijer, A.E.; Middeldorp, J.M. Epstein-Barr Virus mRNA Profiles and Viral DNA Methylation Status in Nasopharyngeal Brushings from Nasopharyngeal Carcinoma Patients Reflect Tumor Origin. Int. J. Cancer 2017, 140, 149–162. [Google Scholar] [CrossRef]
- Martel-Renoir, D.; Grunewald, V.; Touitou, R.; Schwaab, G.; Joab, I. Qualitative Analysis of the Expression of Epstein–Barr Virus Lytic Genes in Nasopharyngeal Carcinoma Biopsies. J. Gen. Virol. 1995, 76, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Li, R.; Chen, M.-Y.; Yu, C.; Tang, L.-Q.; Liu, Y.-M.; Li, J.-P.; Liu, Y.-N.; Luo, Y.-L.; Zhao, Y.; et al. Single-Cell Transcriptomic Analysis Defines the Interplay between Tumor Cells, Viral Infection, and the Microenvironment in Nasopharyngeal Carcinoma. Cell Res. 2020, 30, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Rosemarie, Q.; Sugden, B. Epstein–Barr Virus: How Its Lytic Phase Contributes to Oncogenesis. Microorganisms 2020, 8, 1824. [Google Scholar] [CrossRef] [PubMed]
- Argirion, I.; Pfeiffer, R.M.; Proietti, C.; Coghill, A.E.; Yu, K.J.; Middeldorp, J.M.; Sarathkumara, Y.D.; Hsu, W.-L.; Chien, Y.-C.; Lou, P.-J.; et al. Comparative Analysis of the Humoral Immune Response to the EBV Proteome across EBV-Related Malignancies. Cancer Epidemiol. Biomark. Prev. 2023, 32, 687–696. [Google Scholar] [CrossRef]
- Chia, W.-K.; Teo, M.; Wang, W.-W.; Lee, B.; Ang, S.-F.; Tai, W.-M.; Chee, C.-L.; Ng, J.; Kan, R.; Lim, W.-T.; et al. Adoptive T-Cell Transfer and Chemotherapy in the First-Line Treatment of Metastatic and/or Locally Recurrent Nasopharyngeal Carcinoma. Mol. Ther. 2014, 22, 132–139. [Google Scholar] [CrossRef]
- Li, J.; Chen, Q.-Y.; He, J.; Li, Z.-L.; Tang, X.-F.; Chen, S.-P.; Xie, C.-M.; Li, Y.-Q.; Huang, L.-X.; Ye, S.; et al. Phase I Trial of Adoptively Transferred Tumor-Infiltrating Lymphocyte Immunotherapy Following Concurrent Chemoradiotherapy in Patients with Locoregionally Advanced Nasopharyngeal Carcinoma. OncoImmunology 2015, 4, e976507. [Google Scholar] [CrossRef]
- Lee, S.P. Nasopharyngeal Carcinoma and the EBV-Specific T Cell Response: Prospects for Immunotherapy. Semin. Cancer Biol. 2002, 12, 463–471. [Google Scholar] [CrossRef]
- Gondhowiardjo, S.A.; Adham, M.; Lisnawati; Kodrat, H.; Tobing, D.L.; Handoko; Haryoga, I.M.; Dwiyono, A.G.; Kristian, Y.A. Current Immune-Related Molecular Approach in Combating Nasopharyngeal Cancer. World J. Oncol. 2019, 10, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Fogg, M.; Wirth, L.J.; Daley, H.; Ritz, J.; Posner, M.R.; Wang, F.C.; Lorch, J.H. Epstein-Barr Virus-Specific Adoptive Immunotherapy for Recurrent, Metastatic Nasopharyngeal Carcinoma: Immunotherapy for Nasopharyngeal Cancer. Cancer 2017, 123, 2642–2650. [Google Scholar] [CrossRef]
- Xiao, Z.; Chen, Z. Deciphering Nasopharyngeal Carcinoma Pathogenesis via Proteomics. Expert Rev. Proteom. 2019, 16, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-Y.; Wei, X.-L.; Wang, Y.-Q.; Wang, F.-H. Current Status and Advances of Immunotherapy in Nasopharyngeal Carcinoma. Ther. Adv. Med. Oncol. 2022, 14, 175883592210962. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.T.; Ye, W.; Zeng, Y.-X.; Adami, H.-O. The Evolving Epidemiology of Nasopharyngeal Carcinoma. Cancer Epidemiol. Biomark. Prev. 2021, 30, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Chan, A.T.C.; Cheung, S.-T.; Thomas, W.A.; CroomCarter, D.; Dawson, C.W.; Tsai, C.-H.; Leung, S.-F.; Johnson, P.J.; Huang, D.P. CTL Control of EBV in Nasopharyngeal Carcinoma (NPC): EBV-Specific CTL Responses in the Blood and Tumors of NPC Patients and the Antigen-Processing Function of the Tumor Cells. J. Immunol. 2000, 165, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Liu, J.; Ren, E.C. Structural and Functional Distinctiveness of HLA-A2 Allelic Variants. Immunol. Res. 2012, 53, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.; Burrows, S.R.; Nicholls, J.; Poulsen, L.M. Identification of Cytotoxic T Cell Epitopes within Epstein-Barr Virus (EBV) Oncogene Latent Membrane Protein 1 (LMP1): Evidence for HLA A2 Supertype-Restricted Immune Recognition of EBV-Infected Cells by LMP1-Specific Cytotoxic T Lymphocytes. Eur. J. Immunol. 1998, 28, 451–458. [Google Scholar] [CrossRef]
- Gfeller, D.; Liu, Y.; Racle, J. Contemplating Immunopeptidomes to Better Predict Them. Semin. Immunol. 2023, 66, 101708. [Google Scholar] [CrossRef]
- Peters, B.; Nielsen, M.; Sette, A. T Cell Epitope Predictions. Annu. Rev. Immunol. 2020, 38, 123–145. [Google Scholar] [CrossRef] [PubMed]
- Feola, S.; Chiaro, J.; Cerullo, V. Integrating Immunopeptidome Analysis for the Design and Development of Cancer Vaccines. Semin. Immunol. 2023, 67, 101750. [Google Scholar] [CrossRef]
- Yewdell, J.W. MHC Class I Immunopeptidome: Past, Present, and Future. Mol. Cell. Proteom. 2022, 21, 100230. [Google Scholar] [CrossRef]
- Chong, C.; Coukos, G.; Bassani-Sternberg, M. Identification of Tumor Antigens with Immunopeptidomics. Nat. Biotechnol. 2022, 40, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Mayer, R.L.; Impens, F. Immunopeptidomics for Next-Generation Bacterial Vaccine Development. Trends Microbiol. 2021, 29, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Bassani-Sternberg, M.; Bräunlein, E.; Klar, R.; Engleitner, T.; Sinitcyn, P.; Audehm, S.; Straub, M.; Weber, J.; Slotta-Huspenina, J.; Specht, K.; et al. Direct Identification of Clinically Relevant Neoepitopes Presented on Native Human Melanoma Tissue by Mass Spectrometry. Nat. Commun. 2016, 7, 13404. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, A.M.; Stopfer, L.E.; Ahn, R.; Sanders, E.A.; Sandel, D.A.; Freed-Pastor, W.A.; Rideout, W.M.; Naranjo, S.; Fessenden, T.; Nguyen, K.B.; et al. Deciphering the Immunopeptidome in Vivo Reveals New Tumour Antigens. Nature 2022, 607, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Tso, K.K.-Y.; Yip, K.Y.-L.; Mak, C.K.-Y.; Chung, G.T.-Y.; Lee, S.-D.; Cheung, S.-T.; To, K.-F.; Lo, K.-W. Complete Genomic Sequence of Epstein-Barr Virus in Nasopharyngeal Carcinoma Cell Line C666-1. Infect. Agents Cancer 2013, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Yao, Y.; Chen, H.; Zhang, S.; Cao, S.-M.; Zhang, Z.; Luo, B.; Liu, Z.; Li, Z.; Xiang, T.; et al. Genome Sequencing Analysis Identifies Epstein–Barr Virus Subtypes Associated with High Risk of Nasopharyngeal Carcinoma. Nat. Genet. 2019, 51, 1131–1136. [Google Scholar] [CrossRef]
- Martorelli, D.; Houali, K.; Caggiari, L.; Vaccher, E.; Barzan, L.; Franchin, G.; Gloghini, A.; Pavan, A.; Da Ponte, A.; Tedeschi, R.M.; et al. Spontaneous T Cell Responses to Epstein-Barr Virus-encoded BARF1 Protein and Derived Peptides in Patients with Nasopharyngeal Carcinoma: Bases for Improved Immunotherapy. Int. J. Cancer 2008, 123, 1100–1107. [Google Scholar] [CrossRef]
- Whitney, B.M.; Chan, A.T.C.; Rickinson, A.B.; Lee, S.P.; Lin, C.K.; Johnson, P.J. Frequency of Epstein-Barr Virus-Specific Cytotoxic T Lymphocytes in the Blood of Southern Chinese Blood Donors and Nasopharyngeal Carcinoma Patients. J. Med. Virol. 2002, 67, 359–363. [Google Scholar] [CrossRef]
- Hau, P.M.; Lung, H.L.; Wu, M.; Tsang, C.M.; Wong, K.-L.; Mak, N.K.; Lo, K.W. Targeting Epstein-Barr Virus in Nasopharyngeal Carcinoma. Front. Oncol. 2020, 10, 600. [Google Scholar] [CrossRef]
- Pepperl, S.; Benninger-Döring, G.; Modrow, S.; Wolf, H.; Jilg, W. Immediate-Early Transactivator Rta of Epstein-Barr Virus (EBV) Shows Multiple Epitopes Recognized by EBV-Specific Cytotoxic T Lymphocytes. J. Virol. 1998, 72, 8644–8649. [Google Scholar] [CrossRef]
- Hislop, A.D.; Annels, N.E.; Gudgeon, N.H.; Leese, A.M.; Rickinson, A.B. Epitope-Specific Evolution of Human CD8+ T Cell Responses from Primary to Persistent Phases of Epstein-Barr Virus Infection. J. Exp. Med. 2002, 195, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, V.; Sun, L.; Howard, K.; Sung, S.; Delecluse, H.-J.; Hammerschmidt, W.; Swaminathan, S. Functional Analysis of Epstein-Barr Virus SM Protein: Identification of Amino Acids Essential for Structure, Transactivation, Splicing Inhibition, and Virion Production. J. Virol. 2004, 78, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Murata, T.; Kanda, T.; Isomura, H.; Narita, Y.; Sugimoto, A.; Kawashima, D.; Tsurumi, T. Epstein-Barr Virus Deubiquitinase Downregulates TRAF6-Mediated NF-κB Signaling during Productive Replication. J. Virol. 2013, 87, 4060–4070. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Bongaerts, M.; Hardillo, J.; Tsang, A.; Lo, K.W.; Kortleve, D.; Ma, B.; Debets, R. Transcriptomics of Epstein–Barr Virus Aids to the Classification of T-Cell Evasion in Nasopharyngeal Carcinoma. Curr. Opin. Immunol. 2023, 83, 102335. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.C.; Chen, J.Y.; Liu, M.Y.; Yang, H.I.; Hsu, M.M.; Chen, C.J.; Yang, C.S. Serologic Markers of Epstein-Barr Virus Infection and Nasopharyngeal Carcinoma in Taiwanese Men. N. Engl. J. Med. 2001, 345, 1877–1882. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.F.; Sheng, W.; Cheng, W.M.; Ng, M.H.; Wu, B.H.; Yu, X.; Wei, K.R.; Li, F.G.; Lian, S.F.; Wang, P.P.; et al. Incidence and Mortality of Nasopharyngeal Carcinoma: Interim Analysis of a Cluster Randomized Controlled Screening Trial (PRO-NPC-001) in Southern China. Ann. Oncol. 2019, 30, 1630–1637. [Google Scholar] [CrossRef]
- Yu, X.; Chen, H.; Ji, M. Epstein-Barr Virus-Based Nasopharyngeal Carcinoma Population Screening. Ann. Nasopharynx Cancer 2022, 6, 3. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, X.; Zhang, S.; Duan, X.; Xiang, T.; Zhou, X.; Zhang, W.; Zhang, X.; Feng, Q.; Kang, Y.; et al. T Cell Epitope Screening of Epstein-Barr Virus Fusion Protein gB. J. Virol. 2021, 95, e00081-21. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2021, 50, D543–D552. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | HLA Class I Typing | EBV Strain | Strep-Tagged HLA Overexpression | Lytic Induction |
|---|---|---|---|---|
| CNE2 | A*68:02:01;A*11:01:01 | M81 | A*11:01 | non-lytic |
| 48H | ||||
| A*02:01 | non-lytic | |||
| 48H | ||||
| HK1 | A*24:02:01;A*24:02:01 | Akata | A*11:01 | non-lytic |
| 48H | ||||
| A*02:01 | non-lytic | |||
| 48H | ||||
| C666 | A*24:260;A*24:02:53 | C666-1 | A*11:01 | non-lytic |
| A*02:01 | non-lytic |
| Peptide | Length | Accession | HLA Restriction | EBV Cycle |
|---|---|---|---|---|
| ATIGTAMYK | 9 | EBV-M81-1|BRLF1 | HLA-A*11:01 | Immediate Early Lytic |
| KTLREFFTK | 9 | EBV-M81-1|SMprotein | HLA-A*11:01 | Early Lytic |
| SVGRLTLVNK | 10 | EBV-M81-1|BLLF3 | HLA-A*11:01 | Early Lytic |
| GIAYPPLVH | 9 | EBV-M81-1|BFLF1 | HLA-A*11:01 | Early Lytic |
| ALMPTSGCAQV | 11 | EBV-M81-1|BORF2 | HLA-A*02:01 | Early Lytic |
| GLGEIDWRL | 9 | EBV-M81-1|BKRF3 | HLA-A*02:01 | Early Lytic |
| GLDDRLWAL | 9 | EBV-M81-1|BRRF1 | HLA-A*02:01 | Early Lytic |
| ALWPHPEFL | 9 | EBV-M81-1|BPLF1 | HLA-A*02:01 | Late Lytic |
| AMIEGHLGL | 9 | EBV-Akata|BSLF1 | HLA-A*02:01 | Early Lytic |
| ALERPLAVQL | 10 | EBV-Akata|BMRF1 | HLA-A*02:01 | Early Lytic |
| TLDYKPLSV | 9 | EBV-Akata|BMRF1 | HLA-A*02:01 | Early Lytic |
| RLATVLPGLEV | 11 | EBV-Akata|BALF2 | HLA-A*02:01 | Early Lytic |
| VTFDSPVQR | 9 | EBV-Akata|BBRF1 | HLA-A*11:01 | Late Lytic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, W.; Shi, R.; Luo, Y.; Feng, Z.; Chen, Y.; Zhang, Q.; Zhou, Y.; Liang, J.; Ye, X.; et al. Identification of HLA-A*11:01 and A*02:01-Restricted EBV Peptides Using HLA Peptidomics. Viruses 2024, 16, 669. https://0-doi-org.brum.beds.ac.uk/10.3390/v16050669
Wang Y, Zhang W, Shi R, Luo Y, Feng Z, Chen Y, Zhang Q, Zhou Y, Liang J, Ye X, et al. Identification of HLA-A*11:01 and A*02:01-Restricted EBV Peptides Using HLA Peptidomics. Viruses. 2024; 16(5):669. https://0-doi-org.brum.beds.ac.uk/10.3390/v16050669
Chicago/Turabian StyleWang, Yufei, Wanlin Zhang, Ruona Shi, Yanran Luo, Zhenhuan Feng, Yanhong Chen, Qiuting Zhang, Yan Zhou, Jingtong Liang, Xiaoping Ye, and et al. 2024. "Identification of HLA-A*11:01 and A*02:01-Restricted EBV Peptides Using HLA Peptidomics" Viruses 16, no. 5: 669. https://0-doi-org.brum.beds.ac.uk/10.3390/v16050669