
Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves

College of Horticulture, Nanjing Agricultural University, Nanjing 210095, China
*
Author to whom correspondence should be addressed.
Forests 2024, 15(5), 769; https://0-doi-org.brum.beds.ac.uk/10.3390/f15050769
Submission received: 9 April 2024
/
Revised: 23 April 2024
/
Accepted: 24 April 2024
/
Published: 27 April 2024
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:To provide a theoretical basis for the application of ALA in pear production, the effects of exogenous 5-aminolevulinic acid (ALA) treatment on leaf photosynthetic gas exchange parameters, chlorophyll fast fluorescence properties, and relative expression of the related genes were investigated using pear (Pyrus pyrifolia Nakai cv. ‘Whasan’) as a material in the study. The results show that exogenous ALA treatment improved the photosynthetic gas exchange parameters of pear leaves, upregulated the expression of multiple key genes which are related to ALA biosynthesis, metabolism, and transformation into chlorophylls. GUS staining in tobacco leaves showed that exogenous ALA activated the promoter activity of PypHEMA and PypCHLH genes, implying that the synthesis of endogenous ALA and chlorophylls was promoted by exogenous ALA. Furthermore, ALA promoted the expression of the genes encoding photosystem II (PSII) reaction center proteins, such as core protein D1, inner light-harvesting pigment proteins CP43 and CP47, and cytochrome b559. This led to increased PSII reaction center activity. In addition, ALA alleviated the donor side oxygen-evolving complex inhibition and reduced the closure rate on the receptor side, allowing for increased photochemical electron transfer and reduced heat dissipation while improving the photosynthetic performance index PIabs and PItotal. The findings of this study contribute to a better understanding of ALA’s promotion of plant photosynthetic efficiency, providing valuable insights for further research and potential applications in pear production.
1. Introduction
Pears are the second largest deciduous fruit tree in China after apples, with a total cultivated area of 921.61 × 1000 hm2 in 2021, resulting in a total output 19.88 million tons. Although the pear cultivation area has slightly decreased due to national policy adjustments, the total output continues to grow because of increased unit yield. Cultivation of pear trees is the main income source for many farmers, and the pears are a major fresh fruit consumed by most Chinese residents. However, improper management practices in pear production, such as the overuse of inorganic fertilizers to compensate for insufficient organic fertilizer, can lead to fruit quality issues [1]. As is well known, photosynthesis is a process of matter and energy conversion. It involves the H2O photolysis-induced release of oxygen, conversion of solar energy into electrochemical energy, and the transformation of atmospheric CO2 into carbohydrates with which to store energy in organic compounds. Xu et al. have pointed out that photosynthesis is the material basis and energy source of all life activities in the Earth’s ecosystem [2]. The stronger the photosynthesis of plants, the more organic matter synthesized, the more conducive to the growth and development of plants and the production of higher quality fruits [3]. Therefore, the question of how to improve the photosynthetic performance of plant leaves and how to promote plants to produce more organic matter is a major issue to which producers and researchers pay attention.
5-aminolevulinic acid (ALA) is a key precursor for the biosynthesis of porphyrins such as chlorophylls and heme in all organisms [4]. Recent studies have shown that it is not only a simple biochemical metabolite, but also a multifunctional natural plant growth regulator that can promote seed germination [5,6], seedling growth [6,7], crop yield [8,9,10], and fruit quality [11,12,13,14,15] as well as plant salt tolerance [16], cold tolerance [17,18,19], high temperature tolerance [20] and low light tolerance [17,21,22]. These studies indicate that ALA has important application potential in agricultural and forestry production. In terms of promoting leaf photosynthesis, ALA can significantly increase the chlorophyll content of plants [17,23,24]. At the same time, it can also improve the photochemical efficiency of photosynthetic system II reaction center (PSII-RC) in leaves [25,26], and alleviate photosynthetic system II reaction center (PSI-RC) photosynthetic photoinhibition [12,27]. The effect of ALA on plant photosynthesis is found not only in the normal temperature and light environment, but also when plants encounter different adverse conditions. Recently, Chen et al. proposed that ALA reversed the stomatal closure of apple leaves induced by plant hormone abscisic acid (ABA). They demonstrated that ALA upregulated expressions of protein phosphatase 2A (PP2A) genes, especially the C subunit gene, with a result that the PP2A protein abundance, holoenzyme activity and phosphatase phosphorylation were promoted [28]. The phosphorylated PP2AC interacted with sucrose non-fermenting 1-related protein kinase 2.6 (SnRK2.6), a key component of ABA signaling pathway, causing the latter to become dephosphorylated and block ABA signaling and ABA-induced stomatal closure. Because ABA is a plant stress hormone, which is often accumulated when plants are subjected to stressful conditions, while ALA can reverse the stomatal closure induced by ABA, ALA enables plants to trap more atmospheric CO2 for photosynthetic accumulation, growth, and development of plants under stress [29].
ALA application in pears has been reported to have various effects, such as flower thinning, improved fruit quality, and enhanced anthocyanin accumulation. Firstly, Shen et al. sprayed more than 600 mg L−1 ALA in the blooming period of ‘Hosui’ pear trees and found that it significantly thinned the flowers that opened that day. Consequently, the fruit setting rate was depressed, with increases in fruit size and quality [30]. In this study, the effect of ALA on the promotion of leaf photosynthesis extended at least three months. In another study, when an amino acid foliar fertilizer containing less than 5 mg L−1 ALA was sprayed on pear leaves of 8 cultivars, the leaf chlorophyll content was significantly increased, with higher photosynthetic electron transfer performance [31]. In ‘Akimizu’ pear, a week after ALA spray, the net photosynthetic rate (Pn) and photosynthetic electron transport efficiency were increased, companied with upregulation of the expression of Rubisco (ribulose-1, 5-bisphosphate carboxylase oxygenase) small subunit gene and Rubisco initial activity. Additionally, ALA also increased the activities of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), and ascorbate specific peroxidase (APX), and prevented the accumulation of malondialdehyde (MDA) from membrane lipid peroxidation [32]. Recently, ALA was proposed to alleviate the chilling damage caused to pear flowers by temperatures of −2 °C [33]. In red-skin pears, such as the ‘Yunhongli 2’ [34] and ‘Nanhong’ pears [35], ALA can induce anthocyanin accumulation and promote skin coloring. Despite the potential benefits, the mechanisms behind ALA’s effects on pear biological processes are not yet fully understood.
Recently, our team has reported that ALA promotes peach leaf photosynthesis and photosynthate transport from the source to sink, contributing to high-quality fruit production [15]. Based on this study, we formulated a technical regulation for the application of 5-aminolevulinic acid in peach trees (T/JAASS 88-20 [36]), in which we proposed that ALA should be applied at several specific stages of growth and development of fruit trees. The first is rhizosphere application of ALA solution more than one week before blooming, which can promote floral differentiation and prevent chilling harm from the late spring cold. The second is to spray ALA solution at the late blooming stage to thin flowers, reducing the cost of fruit thinning. The third is to spray ALA solution at the leaves of fruit trees, which can promote cell division of young fruits and leaf development. These three applications of ALA in fruit trees are fundamental for the leaf function and fruit quality. After that, ALA can be applied at 20–30-day intervals until 20–30 days before fruit harvest. However, the procedure has not been tested in the other species of fruit trees. To investigate ALA’s effects on the photosynthetic capacity of pear trees, we conducted a study involving ALA solution application at specific growth stages, including root irrigation before flowering, flower spraying at the end of the full flowering stage, and foliar application at young fruit stage. One week later, we measured the leaf gas exchange, chlorophyll fast fluorescence, expressions of the genes related to chlorophyll synthesis and photosynthetic electron transport carriers. Through some key gene manipulation analyses, we found that exogenous ALA treatment might regulate the biosynthesis of endogenous ALA and chlorophylls. These findings provide a novel insight for ALA application in pear production.
2. Materials and Methods
2.1. Experimental Materials and Treatments
The experiment was conducted in the Jinmei Family Pear Garden, Hushu Street, Nanjing from March to September 2022. This is in the middle and lower reaches of the Yangtze River Plain, with brown–yellow soil, a subtropical monsoon climate, sufficient precipitation, rain and heat in the same season, and four distinct seasons. The 10-year-old pear (Pyrus pyrifolia Nakai cv. ‘Whasan’) trees were planted in the north–south direction, 2 m × 4 m plant spacing, and multi main branches within an open shape. The control and ALA treatment were set up with the trees. A single plant was set as a plot with 10 repeats and random arrangement. The pear trees were treated with ALA solution (provided by Nanjing Hejiachun Biot. Co., Ltd., Nanjing, China) on March 14 (the petal white stage), March 27 (the late blooming stage) and May 8 (six weeks after the fall of flowers). Among these, the treatment on March 14 was pre-flowering root irrigation, the main purpose being to prevent the late spring cold. During the operation, each plant was irrigated with 5 L 10 mg L−1 ALA solution. At the end of the blooming stage on March 27, the same concentration of ALA solution was sprayed on the flowers to remove the late flowering and reduce the fruit thinning cost in the later stage. May 8 was the fruit-set period. One fruit was retained in each inflorescence while the others were artificially thinned. Afterwards, leaves were sprayed with 10 mg L−1 ALA until water dripped from the leaf surface. For each ALA treatment, the control was applied with an equal amount of fresh water. One week after the leaf spraying, the mature functional leaves in the middle of the new peripheral shoots were randomly selected in order to measure the photosynthetic performance and chlorophyll fast fluorescence characteristics on the morning of sunny days. Meanwhile, the leaves were detached and treated with liquid nitrogen, and transported back to the laboratory in ice bottles and stored in a −80 °C refrigerator for subsequent analysis. The field measurements were performed weekly and continued for five weeks.
2.2. Determination of Pigment Content in Leaves
The pigment content of leaves was determined by acetone extraction method [37] and calculated according to the following formulas:
where V is the volume of extracted liquid (mL) and W is the fresh weight of leaves (g). Biological repeats were performed 3 times per treatment.
Chlorophyll a (Chl a) = (12.21 OD663 − 2.81 OD646) × V/1000 W
Chlorophyll b (Chl b) = (20.13 OD646 − 5.03 OD663) × V/1000 W
Carotenoid (Car) = (4.4 OD470–0.01 × Chl a − 0.45 × Chl b) × V/1000 W
The relative chlorophyll content (SPAD) was determined using the SPAD-502PLUS instrument of Konica Minolta, Japan, and the mature leaves were selected in the middle of the peripheral shoots. Each data measurement was repeated 60 times and averaged.
2.3. Determination of Endogenous ALA Content and ALA Biosynthesis and Metabolism in Leaves
According to the method of Harel and Klein [38], the pear leaves (0.1 g) were ground and extracted with 200 mol L−1 acetic acid buffer (pH 4.6). After centrifugation at 5000× g for 15 min, the supernatant was taken and condensed with acetylacetone at 100 °C for 10 min. After cooling to room temperature, 1 mL of newly prepared Ehrlish’s reagent (42 mL glacial acetic acid +8 mL 70% perchloric acid, 1 g dimethylaminobenzaldehyde) was added for color development. After shaking and standing for 10 min, the OD553 value was measured. The standard curve was prepared with ALA standard (Sigma-Aldrich, St. Louis, MO, USA).
To determine the biosynthesis and metabolism of ALA, a certain amount of pear leaves was immersed in a Petri dish containing 20 mmol L−1 levulinic acid (LA, a metabolic inhibitor of ALA) solution, and induced under 240 µmol m−2 s−1 light for 6 h to block ALA metabolism. Then, the accumulated ALA content of leaves was determined according to the method in the above paragraph, which was used as the ALA biosynthesis ability. The difference between the ALA accumulation determined by LA induction and the endogenous ALA content obtained by direct determination was the ALA metabolic capacity.
2.4. Determination of Photosynthetic Gas Exchange Parameters
The gas exchange parameters of pear leaves were measured using a Li-6800 portable photosynthetic system analyzer (Li-COR, Lincoln, NE, USA) between 9:00 and 11:00 on sunny days. The conditions were as follows: leaf chamber temperature 27.0 °C, CO2 concentration 400 μmol mol−1, air humidity 60%, light intensity 1800 μmol m−2 s−1, fan speed 10,000 rpm, gas flow rate 500 μmol s−1, and chamber pressure 0.1 kPa. The instantaneous carboxylation efficiency and water use efficiency were calculated according to the following formulas: instantaneous carboxylation efficiency = Pn/Ci and water use efficiency (WUE) = Pn/Tr. Each treatment was repeated 20 times and the average value was taken.
2.5. Determination of Chlorophyll Fast Fluorescence
Chlorophyll fast fluorescence was measured using the M-PEA multi-function plant efficiency meter (Hansatech, Norfolk, UK). During the determination, the leaves were exposed to saturated pulsed light (3000 μmol m−2 s−1) for 1 s after dark treatment for more than 15 min. The obtained fluorescence induction curves were analyzed by JIP test and MRo/MR according to the methods of Li et al. [39] and Wang et al. [40]. The measurements were repeated 15 times for each treatment and averaged.
2.6. RNA Extraction and RT-qPCR Analysis
According to the kit instructions, the total RNA of pear leaves frozen in liquid nitrogen was extracted by plant RNA extraction kit V1.6 (Biofit, Shanghai, China). The genomic DNA was removed with the Evo M-MLV Reverse Transcription Premix Kit (Accurate, Hunan, China) and reverse transcribed into cDNA. Primers (Table 1) were designed using Primer 5.0 (Premier, Vancouver, Canada). The SYBR Green Pro Taq HS premix qPCR kit (Accurate, Hunan, China) was used to perform qPCR amplification with cDNA as a template and Pyrus bretschneideri actin (LOC103931062) as an internal reference gene. The relative expression levels of genes encoding electron carriers related to chlorophyll synthesis and photosynthetic electron transport were calculated by 2−ΔΔCT method [41]. Each gene was subjected to 3 biological replicates and averaged.
2.7. DNA Extraction and Promoter Sequence Cloning from Plant Materials
The genomic DNA of pear leaves frozen in liquid nitrogen was extracted by Genesand (Beijing, China), an efficient plant genomic DNA extraction kit. The nucleotide sequence of the target gene promoter (about 2 kb upstream the start codon) was retrieved from the National Center for Biotechnology Information (NCBI) database, and specific primers were designed according to the sequence at both ends of the promoter (Table 1). The promoter sequence was cloned using 2 × Phanta Flash Master Mix (P520) high fidelity enzyme (Vazyme, Nanjing, China) using pear genomic DNA as a template. The amplified products were detected by 1.0% agarose gel electrophoresis and the bands containing the target fragment were cut off. The purified bands were then recovered using the DNA gel recovery kit (Genesand, Beijing, China).
2.8. Construction of Plant Gene Expression Vectors
The construction of plant gene expression vectors was undertaken using binary expression vector pBI121. The double enzyme digestion of the carrier was carried out by rapid endonuclease (Transgen, Beijing, China) FlyCut® BamHI (JB101) and FlyCut® HindIII (JB101). After separation by agarose gel electrophoresis, the target fragment was recovered using the DNA gel recovery kit (Genesand, Beijing, China).
The recombination of the target gene promoter and the double enzyme digestion linearized vector was carried out according to the ClonExpress II One Step Cloning Kit (C112) (Vazyme, Nanjing, China) instruction. After ligation, the ligation product was transformed into DH5α competent Escherichia. coli (CAT#: DL1001) (Weidibio, Shanghai, China) at a ratio of 1:10 (v/v) in an ultra-clean bench. After shaking the culture at 37 °C for 6–9 h at 200 rpm until the solution was turbid, the normal E. coli monoclonal colonies were picked and cultured in LB liquid medium (KanR 50 mg/L). PCR reaction was performed using 2 × Rapid Taq Master Mix (Vazyme, Nanjing, China) to detect positive clones. The PCR reaction products were separated by 1% agarose gel electrophoresis, and the bacterial liquid group containing the target band was sent to General Biol. Co., Ltd. (Chuzhou, Anhui, China) for sequencing. The bacterial solution with correct sequencing results was extracted and collected with a high-purity plasmid DNA small extraction kit (Genesand, Beijing, China).
2.9. Agrobacterium Transformation, Tobacco Infection and GUS Qualitative and Quantitative Experiments
According to the requirements of Tsingke (Beijing, China) product specification, the recombinant plasmid was transformed into Agrobacterium GV3101 (TSC-A01). The cells were cultured on a 200 rpm shaker at 28 °C for 24 h until the solution was turbid and inoculated on LB liquid medium containing antibiotics (KanR 50 mg L−1 + Rif 50 mg L−1). The normal Agrobacterium monoclonal colonies were picked, and the Agrobacterium positive clones were detected according to the method in the previous section for subsequent infection.
Using HindIII and BamHI enzymes, the cloned target gene promoter was inserted into the pBI121 vector to obtain a recombinant plasmid. Once confirmed by sequencing, it was transformed into an Agrobacterium (GV3101) competent state in order to obtain the Agrobacterium solution containing the recombinant plasmid. At the same time, the Agrobacterium solution containing the pBI121 empty plasmid, which contained the 35S strong promoter was prepared to produce 35S::GUS. According to the method of Wu et al., the Agrobacterium solution containing different plasmids was injected into the leaves of four-week-old Nicotiana benthamiana, and either water or 1 mg L−1 ALA solution was sprayed. This was repeated three times [42]. After being cultured in the dark for 24 h, the leaves were transferred to the light for 48 h before being collected and the expression level of GUS reporter gene qualitatively and quantitatively detected by the method of Jefferson et al. [43].
In the qualitative detection, the tobacco leaves were completely immersed in the GUS dye solution (X-Gluc mother liquid: X-Gluc base liquid = 1:9) and placed in a constant temperature incubator at 37 °C for 24 h. Then, the leaves were decolored with anhydrous ethanol at 37 °C until they became white, and were then observed and photographed.
In the quantitative detection, tobacco NtActin (LOC107830629) was used as an internal reference. The total RNA of tobacco leaves injected with Agrobacterium was extracted by the method described in 2.6, cDNA was obtained by reverse transcription, and RT-qPCR reaction was performed. The relative expression of GUS gene was calculated by 2−ΔΔCT method [41]. Each treatment was performed with three biological replicates. Primer names and sequences are shown in Table 1.
2.10. Data Analysis
All data were calculated and processed using Microsoft Excel. The charts were drawn using GraphPad Prism 9.0.1, and Photoshop CC (2015) was used to assist modification. SPSS 17.0 was used for statistical analysis. Two-way analysis of variance (F-test) and Duncan’s test were used to analyze the different times after ALA treatment and the significant effect of ALA treatment (* p < 0.05, ** p < 0.01).
3. Results
3.1. Effects of Exogenous ALA Treatment on Photosynthetic Gas Exchange Parameters in Pear Leaves
Figure 1 shows the changes of photosynthetic gas exchange parameters in pear leaves over five weeks (May 15 to June 12) after exogenous ALA foliar application. Regardless of control or treatment, the net photosynthetic rate (Pn) of pear leaves was stable and had an upward trend overall, indicating that the photosynthetic performance of pear leaves was stable and gradually increased at this stage. However, in week 2 (May 22), the Pn of both leaves decreased. This may be the result of high temperature stress caused by a sudden rise in temperature, from the 30–34 °C recorded in the other weeks to 39 °C. The Pn of leaves in ALA treatment was significantly higher than that of control, which lasted for at least five weeks. During this period, ALA-induced Pn increased by 29.28% (p < 0.01, n = 100).
The transpiration rate (Tr) of pear leaves one week after ALA spraying was significantly higher than that of the control, but was significantly decreased at the second week while that of the control was increased slightly. After the third week, Tr of the treated leaves was gradually higher than that of control. At five weeks, the Tr of ALA treated leaves was 13.94% higher than that of control (p < 0.01, n = 100). Similarly, the stomatal conductance (Gsw) of the pear leaves treated with ALA was also decreased significantly at the second week after ALA treatment. Overall, however, the Gsw of leaves treated with ALA was 18.73% higher than that of control (p < 0.01, n = 100). In contrast, there was no overall significant difference in CO2 concentration between pear leaves due to ALA (p > 0.05), though the instantaneous carboxylation efficiency (Pn/Ci) of pear leaves was significantly increased by 32.30% (p < 0.01) due to ALA treatment. Water use efficiency in ALA treatment was also increased by 11.16% (p < 0.01). In conclusion, ALA treatment significantly improved the photosynthetic performance of pear leaves.
3.2. Effects of Exogenous ALA Treatment on Chlorophyll Fluorescence Characteristics of Pear Leaves
Figure 2A–E show the effects of exogenous ALA on the chlorophyll fast fluorescence induction kinetic curves of pear leaves. After the pear leaves were darkened for 15 min and then irradiated with strong light, the fluorescence intensity rapidly increased from O point (0 ms) to J point (3 ms). At this time, the growth rate slowed slightly, and then increased rapidly again. After I point (30 ms), it gradually reached the highest point P (t ≈ 300 ms), and then stabilized, showing a typical OJIP curve. Within 1–5 weeks of the foliar spraying of ALA solution, the initial fluorescence (Fo) of the treated leaves decreased slightly, while the maximum fluorescence yield (Fm) at P point increased significantly, resulting in a significant increase in Fv and photochemical efficiency (Fv/Fm ≡ φPo).
Figure 2F–J shows the 820 nm reflection fluorescence absorption curve of pear leaves within five weeks of the foliar spraying of ALA. The decrease stage of MRo/MR represents the oxidation activity of PSI-RC, and the increase stage represents the reduction process of PSI-RC. The MRo/MR ratio began to decrease from 0.7 ms (JIP time) and reached its lowest value between t = 10–30 ms. Then, PSI-RC was gradually reduced by the electrons transferred from PSII-RC, so that the reflected fluorescence gradually increased and reached its peak at about 300 ms. At this time, PSI-RC was completely restored. The results show that the MRo/MR of the treated leaves was slightly lower than that of the control at 2–4 weeks after ALA treatment, and that the difference was significant at p = 0.05 at the second week. However, it was not significant at other times (p > 0.05), indicating that the promotion of ALA treatment on PSI-RC activity in pear leaves appeared two weeks after ALA treatment, but soon disappeared. In addition, among the weekly MRo/MR minimum, the first week was the lowest, and then gradually increased until, in the fourth week, it reached its maximum. There was no difference between the other three weeks. This result indicates that the redox activity of PSI-RC decreased from May 15 to June 12 during the development of pear leaves.
The chlorophyll fluorescence parameters calculated according to the chlorophyll fluorescence curve in Figure 2 are shown in Table 2. Wk represents the degree of inhibition of the PSII-RC donor-side oxygen-evolving complex (OEC). The Wk of ALA-treated leaves was significantly lower than that of the control in the first three weeks (p < 0.05), indicating that ALA can increase OEC activity and reduce the inhibitory effect of internal and external factors on the OEC. Mo is the maximum rate of QA reduction on the PSII-RC receptor side. The larger the value, the faster the PSII-RC is closed. Within three weeks of ALA treatment, the Mo of the treated leaves was significantly lower than that of the control (p < 0.05), indicating that ALA can delay the closure of PSII-RC and facilitate the transfer of photosynthetic electrons from PSII to QA−. Sm represents the energy required for QA to be completely reduced, reflecting the capacity of the PQ reservoir on the PSII-RC receptor side. From Table 2, the Sm of the first three weeks after ALA treatment were significantly higher than those of the control (p < 0.05), indicating that ALA increased PQ storage capacity. Φo and φPo represent the probability of the trapped excitons transferring electrons to other electron acceptors downstream of the electron transport chain QA- and the maximum photochemical efficiency of PSII, respectively. Within five weeks of ALA treatment, the φo and φPo in the ALA treatment were significantly higher than those in the control (p < 0.05). Similarly, φEo, the quantum yield of light energy absorbed by PSII-RC for electron transfer after ALA treatment was significantly higher than that of the control within five weeks, while φDo, the quantum yield for heat dissipation was significantly lower than that of the control (p < 0.05). RC/CS represents the density of active PSII reaction centers per unit area, while PIABS and PItotal represent the photosynthetic performance index based on light energy absorption and the photosynthetic performance index including PSI and PSII, respectively. The RC/CS of ALA-treated leaves was significantly higher than that of the control within five weeks, the PIabs was significantly higher than that of the control at three weeks, and the PItotal was significantly higher than that of the control within four weeks (p < 0.05). In summary, the promotion of ALA on the photosynthetic electron transport performance of pear leaves can be maintained for at least three weeks, and some parameters can be maintained for five weeks. In addition, VPSI is the maximum slope of the descending section of the 820 nm modulated reflection fluorescence curve (t = 0.7 to 3 ms in this study). This reflects the maximum rate of oxidation of the PSI reaction center. ALA treatment had no significant effect on the VPSI of pear leaves, but significantly increased the VPSII-PSI of pear leaves (p < 0.01). This indicates that ALA had no effect on the PSI reaction center of pear leaves, but that it promoted the transfer of PSII-RC electrons to PSI-RC and the reduction of PSI-RC.
3.3. Effects of Exogenous ALA Treatment on Endogenous ALA Biosynthesis and Metabolism in Pear Leaves
Figure 3 shows that the endogenous ALA content and the synthesis ability of pear leaves increased first and then decreased during the test period, while the ALA metabolic decomposition ability was at a low level except for May 15. Exogenous ALA treatment significantly increased endogenous ALA content in pear leaves. This difference appeared from the first week of leaf spraying and remained until the fifth week. The endogenous ALA content induced by ALA increased by 41.99% (p < 0.01) within five weeks. When the isolated pear leaves were treated with levulinic acid and then exposed to light, the endogenous ALA biosynthesis was normal but the metabolism was blocked. In this way, the ALA synthesis ability of pear leaves can be assessed, with the difference between ALA synthesis and the endogenous content being the amount of ALA metabolic decomposition. From Figure 3B, the endogenous ALA synthesis ability of pear leaves increased first and then decreased from May 15 to June 12. This trend is consistent with the endogenous ALA content. Exogenous ALA treatment significantly improved the ability of endogenous ALA synthesis. The mean value within five weeks was increased by 54.54% (p < 0.01). Similarly, exogenous ALA treatment promoted the metabolic decomposition of endogenous ALA in pear leaves (p < 0.01), but this effect was mainly reflected in the second week after ALA treatment. At other time points, although there were differences, they were not statistically significant. This means that spraying pear leaves with exogenous ALA in early May not only promotes endogenous ALA synthesis, but also promotes ALA metabolism. The promotion of exogenous ALA on endogenous ALA synthesis was maintained for at least five weeks, but the promoting effect on ALA metabolism was found mainly in the second week after treatment.
3.4. Effects of Exogenous ALA Treatment on Photosynthetic Pigment Content in Pear Leaves
Figure 4 shows the effect of exogenous ALA treatment on photosynthetic pigment content in pear leaves. Between May 15 and June 12, the photosynthetic pigments of pear leaves increased first and then decreased. One can see, from the results of the relative chlorophyll content (SPAD) (Figure 4A), that, one week after ALA treatment, the SPAD of the treated leaves was slightly higher than that of the control but not within significance (p > 0.05). Thereafter, it was significantly higher than the control. Figure 4B–D show that the chlorophyll a, chlorophyll b and total chlorophyll content of pear leaves after one week of ALA treatment were significantly higher than those of the control. Although there was no difference in Chl b content between the two groups at the second week, the differences at other time points were significant p = 0.05, indicating that ALA treatment promoted chlorophyll accumulation in pear leaves and alleviated the decline.
Specifically, the average values of Chl a, Chl b and total chlorophyll in ALA-treated leaves within five weeks were 11.38%, 8.59% and 10.44% higher than those in the control, respectively (p < 0.01). In addition, ALA was seen to promote the increase of Chl a/b ratio in pear leaves (p < 0.01), indicating that ALA promoted the accumulation of chlorophyll a in leaves, but was not conducive to the conversion of Chl a into Chl b. Similarly, the carotenoid content of pear leaves also showed a trend of ‘first rise and then fall’ within five weeks after treatment, and ALA treatment promoted the increase of Car content in leaves (Figure 4F). The five-week average was 16.25% higher than the control (p < 0.01).
3.5. Effects of Exogenous ALA Treatment on the Gene Expressions Related with Chlorophyll Synthesis
To explore the mechanism by which exogenous ALA increases chlorophyll content in pear leaves, we analyzed the relative expression of the primary genes related to ALA and chlorophyll biosynthesis after the spraying of ALA. With the exception of encoding porphobilinogen deaminase (HEMC) and encoding uroporphyrinogen III decarboxylase (HEME), which were not detected, the relative expressions of other genes are shown in Figure 5. The relative expressions of two key genes, HEMA and GSA, which are involved in ALA synthesis, were significantly upregulated after exogenous ALA treatment. Among these, HEMA expression had the largest increase in the first week after ALA treatment, and was still significantly higher than the control in the second and third weeks; however, in the fourth and fifth week, it was significantly lower than the control. GSA expression was also significantly higher than that of the control from the first week to the fourth week after ALA treatment, although it was significantly lower than that of the control at the fifth week. The expression of HEMB, which encodes the ALA dehydrase and catalyzes the ALA metabolism to produce porphobilinogen, was significantly higher than the control at the second and fourth week after ALA treatment, but no significant difference was found at other times. The expression of HEMD, which encodes the enzyme catalyzing the formation of uroporphyrinogen III from hydroxymethylbilane, was significantly lower than that of the control in the first two weeks of ALA treatment, nevertheless, it was significantly higher than the control at the third and fourth weeks. Overall, ALA treatment downregulated the gene expression (p < 0.05). The enzyme encoded by HEMF catalyzes the conversion of coproporphyrinogen III to protoporphyrin IX. From Figure 5, the expression of the HEMF gene in ALA-treated leaves was significantly higher than that in the control, at least within four weeks (p < 0.05). A similar situation occurred in HEMG (encoding the enzyme that catalyzes protoporphyrinogen IX to protoporphyrin IX).
The CHLH encoding enzyme catalyzes the chelation of protoporphyrin IX with Mg2+ to produce Mg-protoporphyrin IX. This is the first gene to control the branch of chlorophyll biosynthesis. From Figure 5, the relative expression of CHLH in leaves treated with ALA was significantly higher than that of control at week 1, which lasted up to three weeks. Thereafter, the gene expressions were lower than controls. The CHLM encoding enzyme catalyzes the formation of Mg-protoporphyrin IX into Mg-protoporphyrin IX methyl ester. The relative expression of this gene was significantly higher than that of the control at the third and fourth week after ALA treatment but was lower than that of the control at other time points. Overall, ALA treatment downregulated the expression of this gene (p < 0.05). The CRD1 encoding enzyme catalyzes the conversion of Mg-protoporphyrin IX methyl ester to divinyl protochlorophyllide. ALA treatment promoted the upregulation of CRD1 expression, which was manifested in the 2–4 weeks after treatment (p < 0.05). The DVR gene is different, the encoding enzyme catalyzes the conversion of divinyl protochlorophyllide to pchlide and was significantly upregulated at the second and fourth weeks after ALA treatment; however, overall, ALA treatment downregulated DVR gene expression (p < 0.05). The enzyme encoded by POR catalyzes the conversion of pchlide into chlide. At the second and third weeks after ALA treatment, the expression of POR was significantly higher than that of the control. The enzyme encoded by CHLG catalyzes the conversion of chlide into chlorophyll a. Its expression profile is like that of DVR, which was significantly upregulated at 3–4 weeks after ALA treatment, but lower than that of the control at other times. Overall, the gene expression was downregulated by ALA (p < 0.05). The enzyme encoded by CAO catalyzes the conversion of chlorophyll a into chlorophyll b. After ALA treatment, the relative expression of CAO was lower than that of the control (p < 0.05), indicating that ALA treatment inhibited the conversion of chlorophyll a to chlorophyll b.
Overall, ALA promoted the expression of genes encoding its own synthesis, metabolism, and transformation into chlorophyll. The effects were significant within four weeks after ALA treatment, the expression of most genes was upregulated to varying degrees.
3.6. Effect of Exogenous ALA Treatment on Promoter Activity of Genes Related to ALA and Chlorophyll Synthesis
To further explore the mechanism by which ALA regulates chlorophyll synthesis, we selected five genes, HEMA, GSA, HEMF, HEMG and CHLH, which were continuously upregulated from the first week after ALA treatment, in order to analyze the activation of ALA on the promoters of these genes by GUS staining.
The structural map of the pBI121 recombinant vectors carrying the promoters of the target genes was shown in Figure 6A. The Agrobacterium solution carrying the recombinant plasmid was injected into tobacco leaves, and some leaves were selected to spray 1 mg L−1 exogenous ALA solution. Then, the tobacco plants were cultured under dark conditions for 24 h, and transferred to light for 48 h. Leaves were collected and GUS staining was performed with the results in Figure 6B (HEMG gene expression vector was not successfully constructed). The GUS staining degree of tobacco leaves injected with the 35S::GUS empty vector was highest, while those of proPypHEMA::GUS and proPypCHLH::GUS after ALA treatment were significantly higher than that of the control; however, those of proPypGSA::GUS and proPypHEMF::GUS were not significantly stained with the control. Figure 6C shows the relative expression of the GUS gene in tobacco leaves after transient infection. The relative expressions of the GUS gene in tobacco leaves injected with proPypHEMA::GUS and proPypCHLH::GUS were significantly higher than that of the control after ALA treatment, and significantly lower than that of the leaves injected with the empty vector. The relative expressions of the GUS gene in the leaves treated with proPypGSA::GUS and proPypHEMF::GUS bacterial solution injection and the control were not significantly different. Therefore, exogenous ALA treatment may promote gene expression by upregulating the promoter activity of endogenous ALA synthesis key gene HEMA and chlorophyll synthesis key gene CHLH, thus leading to the increase of endogenous ALA and chlorophyll content in pear leaves.
3.7. Expression Analysis of PSII-Related Genes in Pear Leaves Treated with Exogenous ALA
From the results of the chlorophyll rapid fluorescence curve, the promoting effect of ALA on photosynthetic electron transport in pear leaves is mainly manifested in PSII-RC. To explore the mechanism, we further measured the relative expressions of the genes encoding the main members of PSII-RC in pear leaves within five weeks of ALA treatment. From Figure 7, the genes PsbB and PsbC encoding the main members of the inner light-harvesting pigment protein complex CP43 and CP47 were significantly up-regulated at one week after ALA treatment, but this effect disappeared at two weeks. The gene PsbA encoding PSII-RC core protein D1 was significantly higher than that of the control after two weeks of ALA treatment. Similarly, ALA continuously promoted the expressions of PsbE and PsbF genes encoding the Cyt b559 branch core protein for at least four weeks. In addition, PsbY, Psb27, psbH, PsbJ and PsbL were upregulated to varying degrees in the second week after ALA treatment, where PsbY is related to the photoprotection of Cyt b559. Psb27 and psbH are involved in the assembly of PSII-RC by binding to antenna proteins CP43 and CP47, and PsbJ and PsbL may be involved in the light energy conversion process. However, there were no obvious regular changes in the expression of other genes.
The above results indicate that ALA treatment may maintain the stability of the PSII reaction center and promote plant photosynthesis by promoting the expression of genes encoding PSII-RC core structural proteins and other functional small molecule proteins.
3.8. Effect of Exogenous ALA Treatment on Promoter Activity of PSII-Related Genes
To further explore the mechanism of ALA regulating the PSII reaction center protein, nine genes whose expressions were upregulated in the first two weeks after ALA treatment, including PsbA, PsbB, PsbC, PsbE, PsbF, PsbH, PsbJ, PsbL, and Psb27, were selected in this study. GUS staining was used to analyze the activation effect of ALA qualitatively and quantitatively on the promoters of these genes.
During the vector construction, only PsbA, PsbB and PsbJ were successfully constructed. The structural map of the pBI121 recombinant vectors carrying the promoters of the target genes is shown in Figure 8A. The Agrobacterium solution carrying the recombinant plasmid was injected into tobacco leaves, and some leaves were selected to be sprayed with 1 mg L−1 exogenous ALA solution. Then, the tobacco plants were cultured under dark conditions for 24 h and transferred to light conditions for 48 h. Leaves were collected and GUS staining was performed. The results (Figure 8B) show that the GUS staining degree of tobacco leaves injected with the 35S::GUS empty vector bacterial solution was the highest, while that with ALA-treated bacterial solution was not significantly different from that of the control. Figure 8C shows that there was no significant difference in the relative expressions of the GUS gene promoted by proPypPsbA::GUS, proPypPsbB::GUS and proPypPsbJ::GUS in either the ALA treatment or the control. This indicates that exogenous ALA has no significant effect on the promoter activity of PSII reaction center protein genes of pear leaves.
4. Discussion
As early as 1997, Hotta et al. had proposed that exogenous ALA improves the photosynthetic efficiency of leaves of various plants [44]. Over the decades, the role of ALA in promoting photosynthesis has been confirmed in apple [11], grape [12], peach [15], melon [17], cucumber [24], pear [30], strawberry [45], and wheat [46]. At present, ALA’s promotional effects on leaf photosynthesis have been widely utilized in various agricultural and forestry plant productions [47]. In peach, we have developed the “technical regulations for the application of 5-aminolevulinic acid to peach trees” (TJAASS 88-2023 [36]), but whether this is suitable for pear production has not been proven. In this study, we applied ALA to the roots of pear trees before flowering, then sprayed the flowers at the late flowering stage, and sprayed the leaves at the young fruit development period, according to the regulation in peach. After that, we measured the photosynthetic gas exchange parameters on a weekly basis and the chlorophyll rapid fluorescence and gene expressions associated with chlorophyll synthesis and PSII-RC member proteins over the following five weeks, the results provide a theoretical basis for the elucidation of the mechanism of ALA in improving the photosynthetic efficiency in pear leaves.
Firstly, the Pn of pear leaves was significantly increased within five weeks after foliar spraying of ALA (Figure 1A). At the same time, the change trend of leaf Gsw was highly similar to that of Pn (Figure 1C). This is consistent with the results of Zhao et al. (2014) which state that ALA improves the photosynthetic capacity of tomato leaves under salt stress by maintaining the normal opening of stomata and reducing stomatal limitation [26]. In apple, ALA has been shown to upregulate PP2AC gene expression and PP2A protein phosphatase activity, promote SnRK2.6 dephosphorylation [28,29], downregulate Ca2+ and H2O2 content in guard cells [48], and upregulate flavonol content [49], thereby blocking ABA signaling and reversing ABA-induced stomatal closure [50]. ALA has a significant effect on the promotion of stomatal opening in pear leaves (p < 0.05), although the mechanism in pear leaves needs to be clarified further. In grape, exogenous ALA can alleviate ABA-induced stomatal closure when seedlings are subjected to drought stress [51]. However, compared with stomatal aperture, the promotion of ALA on the carboxylation efficiency of pear leaves may be more important. Within five weeks of ALA treatment, the instantaneous carboxylation efficiency of leaves remained at a high level, and the average Pn/Ci was 32.3% higher than that of the control, which was much higher than the average increase of Gsw (18.73%). In addition, ALA treatment significantly increased the water use efficiency of pear leaves (Figure 1F). These results suggest that the promotion of ALA on the photosynthetic efficiency of pear leaves has a variety of performances, and that this promotion is a comprehensive result of multiple factors.
Secondly, during the field experiment, the maximum temperatures were generally maintained at 30–34 °C. However, in the second week (May 22), the temperature soared to 39 °C. This may be an important reason for the decrease of Pn at this time point. From Figure 1, Pn, Tr and Gsw decreased significantly in the second week. However, the Pn and Gsw of leaves after ALA treatment were still significantly higher than those of the control, indicating that exogenous ALA improved the high temperature resistance of pear. This is similar to the observation shown in [20], where ALA was reported to increase leaf gas exchange parameters and photochemical efficiency during high temperature condition.
Thirdly, chlorophylls are the main pigment with which plant chloroplasts capture solar energy and drive electrons to the photosynthetic reaction center [52]. It has long been known that ALA is a key precursor of chlorophyll biosynthesis [53,54], and exogenous ALA treatment can promote plant chlorophyll synthesis [24,55]. Moreover, ALA can be used as a regulator for chlorophyll synthesis. Tanaka et al. (1993) have proposed that ALA promotes the conversion of chlorophyll a into chlorophyll b in cucumber leaves, which bind to the light-harvesting pigment protein complex II (LHCII) apoprotein to avoid the hydrolysis activity of chlorophyllase to Chl a, thereby increasing chlorophyll content [53]. The authors proposed that the Chl b/a ratio of leaves was increased after ALA treatment. However, we did not find a similar phenomenon in this study. Foliar application of ALA not only promoted the increase of Chl a, Chl b content and total chlorophyll content in pear leaves, but also significantly increased Chl a/b ratio. Therefore, different species may have different responses to ALA treatment in the conversion of chlorophylls. In addition, ALA promoted the increase of carotenoid content in pear leaves (Figure 4F). This is similar to tomato fruit [56], but opposite to tomato leaves [57]. Therefore, the relationship between ALA and carotenoids needs to be further studied.
Wu et al. (2018) have proposed that exogenous ALA upregulates gene expressions such as HEMA (encoding the key enzyme GluTR), CHLH (encoding chlorophyll chelatase), and POR (encoding protochlorophyllate oxidoreductase) in cucumber leaves under salt stress [24]. Wang et al. (2021) have also proposed that ALA promotes the expression of HEMA1 and HEMB in pepper leaves under low temperature stress [58]. These indicate that exogenous ALA promotes endogenous ALA synthesis and metabolic decomposition. The results in the current study show that ALA not only promotes the expression of HEMA, GSA and HEMB, three key genes controlling ALA synthesis and metabolism (Figure 5), but also promote the synthesis and metabolism of endogenous ALA in pear leaves. Nevertheless, because exogenous ALA promotes the increase of endogenous ALA content, ALA promotion of the biosynthesis is greater than of the metabolism (Figure 3). The GUS staining has shown that ALA promotes the promoter activity of PypHEMA (Figure 6C). This not only validates the results of previous studies in cucumber [24] and pepper [58], but also proves that exogenous ALA itself induces biosynthesis. Thus, exogenous ALA treatment can exhibit these biological functions over a long period, and thus need not be applied too frequently. In ‘Housui’ pear, the increased photochemical efficiency induced by ALA application at the end of full blooming can be observed after 98 days [30]. The reason for this has never been clear. The finding in the current study, that exogenous ALA upregulates the gene promoter activity of endogenous ALA synthesis, can well explain the long-term effectiveness of ALA on leaf photosynthesis. However, the promoter of GSA, another key gene for endogenous ALA biosynthesis, was not regulated by ALA (Figure 6C). In addition, the mechanism of exogenous ALA activating HEMA promoter remains to be studied further.
Fourthly, chlorophyll synthesis is not only affected by ALA synthesis and metabolism, but also regulated by many other genes. In grape, Yang et al. (2023) have proposed that exogenous ALA upregulates the expressions of DVR, POR, CHLG and other genes, thereby promoting the increases of the leaf chlorophyll content [51]. In the current study, gene expressions related to chlorophyll synthesis were detected in pear leaves within five weeks after exogenous ALA treatment. The results show that the relative expressions of five genes, HEMD, CHLM, DVR, CHLG and CAO, were significantly lower than those of the control. The relative expressions of other genes, including HEMF, HEMG, CHLH, CRD1 and POR, were significantly higher than those of the control (Figure 5). These show that exogenous ALA treatment can promote the expressions of chlorophyll synthesis genes in pear leaves to varying degrees. GUS staining showed that exogenous ALA stimulated the activity of the PypCHLH promoter (Figure 6C). CHLH encodes the H subunit of Mg-chelating enzyme, which catalyzes the chelation of Mg2+ into the porphyrin ring to form Mg-protoporphyrin IX. This is the first gene in the branch of chlorophyll synthesis [59]. The activation of ALA on the gene promoter may be an important mechanism for the upregulation of chlorophyll content in pear leaves. On the other hand, ALA treatment downregulates CAO expression in pear leaves. This may be the main reason for the increase of Chl a/b.
Fifthly, we detected the chlorophyll fast fluorescence curve and 820 nm modulated reflection fluorescence curve of pear leaves within five weeks of ALA treatment and analyzed the effects of ALA on the activity of PSII-RC and PSI-RC in pear leaves using fluorescence data. As shown in Figure 2 and Table 2, the promotion of ALA on PSII-RC activity in pear leaves was significantly higher than that of PSI-RC. VPSI reflects the electron loss rate in the PSI-RC because of photo oxidization after strong light irradiation. We measured these for five weeks and, without exception, did not observe the effect of ALA. On the contrary, VPSII-PSI reflects the ability of electrons transferred from PSII-RC to reduce PSI-RC [40]. The VPSII-PSI of pear leaves after ALA treatment was always found to be higher than that of the control, and the difference was found to be significant in the second week (p < 0.05). Moreover, Wk is a parameter reflecting the inhibition of the PSII-RC donor side. φPo reflects PSII-RC activity. Mo, φo and Sm reflect the characteristics of the receptor side of PSII-RC. We found that ALA treatment reduced Wk and Mo, but increased φPo, φo and Sm. These effects were significant within at least three weeks of ALA treatment. The quantum yield, φEo, and the active reaction center density RC/CS for photosynthetic electron transport in pear leaves were significantly higher than those in the control, while the quantum yield for heat dissipation (φDo) was lower than that in the control. The photosynthetic performance index, PIabs, of ALA-treated leaves was significantly higher than that of the control. In addition, PItotal, including the two photosynthetic reaction centers of PSII and PSI, also increased significantly due to ALA treatment (Table 2). This indicates that ALA also has a promoting effect on the activity of the PSI reaction center. From Figure 2, one can see that the minimum value of MRo/MR of 820 nm modulated reflectance fluorescence curve of pear leaves treated with ALA was lower than that of the control within 2–4 weeks, indicating that the ability of the PSI reaction center to reduce terminal electron acceptors was higher than that of the control [60]. In strawberry, Sun et al. (2011) have proposed that the promotion of ALA on photosynthetic efficiency is related to the activity of PSII-RC and PSI-RC [61]. In pear, Shen et al. (2011) have suggested that ALA has a significant promoting effect on the PSII reaction center of sand pear leaves [31]. Therefore, the regulation mechanism of ALA on PSII-RC activity in pear leaves is worthy of further study, and the effect of ALA on PSI-RC cannot be ignored.
Sixthly, in view of the important promoting effect of ALA on PSII-RC in pear leaves, the gene expressions of PSII-RC-related proteins to ALA treatment were determined in this study. The results show that the expressions of genes encoding the PSII-RC core protein D1 (PsbA), the inner light-harvesting pigment proteins CP43 (PsbB) and the CP47 (PsbC), cytochrome b559 (PsbE and PsbF) were significantly upregulated in pear leaves after spraying of ALA solution for one week. D1 is the core structure of PSII-RC [62] and the most vulnerable component of PSII [63]. ALA treatment significantly increased PsbA gene expression and D1 protein content in wheat under drought stress [46]. CP43 and CP47 connect PSII-RC and LHCII to transfer the energy generated by excitation to the reaction center [64]. As an electron transport branch, Cyt b559 has a light protective effect [65]. In addition, the genes encoding PSII-RC small molecule proteins, such as PsbY, Psb27, PsbH, PsbJ and PsbL were found to be significantly higher than those of the control at 2–4 weeks of ALA treatment (Figure 7). Therefore, it is suggested that the promoting effect of ALA on the photosynthesis of pear leaves may play a protective and promoting role by promoting the expression of PSII-RC core structural proteins and the coding genes of various small molecule proteins, and that this promoting effect has no obvious relationship with the regulation of gene transcription initiation. However, we did not find any direct regulation of ALA on the gene expressions, which needs further study.
5. Conclusions
In summary, the promotion of ALA on the photosynthesis of pear leaves can be maintained for at least three weeks, which means that ALA application is not frequently needed. Conversely, ALA should be applied at different specific stages. At the early stage of plant growth, rhizosphere application to prevent late spring coldness before fruit tree blooming, flower spraying to thin surplus flowers at the end of full blooming and foliar application to promote leaf development are all important for leaf photosynthesis and fruit quality. In the process of chlorophyll synthesis, exogenous ALA promotes its own synthesis and metabolism, promotes the branch of chlorophyll synthesis, and increases the content of photosynthetic pigments. In the process of photosynthetic electron transport, ALA promotes the expression of PSII-RC core protein genes in pear leaves with higher PSII-RC activity at the donor side and the accepting side, which facilitates the promotion of photosynthetic electron transport and energy conversion. Our published data suggest that the single fruit weight and the soluble sugar content of ALA treatment were, respectively, 19.7% and 14.5% higher than those of the control group when fruits were mature (p < 0.05), which may be the results of the increased photosynthesis. However, the regulatory mechanisms of ALA on photosynthetic dark reaction (such as RuBP carboxylase activity) of pear remains to be further studied.
Author Contributions
Conceptualization, X.W. and L.W.; experimental, X.W. and J.Z.; formal analysis, X.W. and J.Z.; writing—original draft preparation, X.W. and L.W.; writing—review and editing, X.W. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32230097, 31772253), the Jiangsu Special Fund for Frontier Foundation Research of Carbon Peaking and Carbon Neutralization (BK20220005), the Jiangsu Agricultural Science and Technology Innovation Fund (CX(20)2023), the Fundamental Research Funds for the Central Universities (YDZX2023018).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Thanks to Yuyan An, Yan Zhong for their guidance to experiment and Longbo Liu, Xing Gan, Ruolin Liang, and Shen Zhang for their contributions to sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, Z.; Zhang, J.; Chen, Z.G. Analysis of the value chain of China’s pear fruit industry. Chin. Rural. Econ. 2008, 1, 63–72. [Google Scholar]
- Xu, D. Photosynthetic rate, photosynthetic efficiency, and crop yield. Bull. Biol. 1999, 1, 11–13. [Google Scholar]
- Peng, L.L.; Jiang, W.B.; Han, J. Effects of source-sink relationship change on yield and quality in fruit tree. Nonwood For. Res. 2012, 30, 134–140. [Google Scholar]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef]
- Wang, L.J.; Jiang, W.B.; Liu, H.; Liu, W.Q.; Kagn, L.; Hou, X.L. Promotion by 5-aminolevulinic acid of germination of pakchoi (Brassica campestris ssp. chinensis var. communis Tsen et Lee) seeds under salt stress. J. Integra Plant Biol. 2005, 47, 1084–1091. [Google Scholar] [CrossRef]
- Lukšienė, Ž.; Danilčenko, H.; Tarasevičienė, Ž.; Anusevičius, Ž.; Marozienė, A.; Nivinskas, H. New approach to the fungal decontamination of wheat used for wheat sprouts: Effects of aminolevulinic acid. Int. J. Food Microbiol. 2007, 116, 153–158. [Google Scholar] [CrossRef]
- Mai, H.X.; Wang, H.X.; Hu, Y.P.; Han, W.Y.; Li, Z.M.; Pang, Z.P.; Zhou, H.G. Effects of growth regulator ALA on growth and physiology traits of potted lily. Jiangsu Agric. Sci. 2021, 49, 9–103. [Google Scholar]
- Kong, L.G.; Wang, Y.H.; Han, X.D.; Bo, H.X.; Xun, J.; Wang, L.J. Effect of Cropspring foliar fertilizer on growth and quality of rice under different nitrogen supplies. Jiangsu J. Agric. Sci. 2018, 34, 90–798. [Google Scholar]
- Li, M.A.; Ma, L.; Hao, Q.; An, Y.Y.; Wang, L.J. Effect of 5-aminolevulinic acid on leaf photosynthetic characteristics, yield, and quality of potato. China Veg. 2020, 1, 43–52. [Google Scholar]
- Sheng, Y.F.; Yang, L.L.; Wang, J.K.; Chen, Y. Effects of 5-aminolevulinic acid on yield and benefit of carrot. Hortic. Seed 2020, 40, 8–9+11. [Google Scholar]
- Gao, J.J.; Feng, X.X.; Duan, C.H.; Li, J.H.; Shi, Z.X.; Gao, C.Y.; Wang, L.J. Effects of 5-aminolevulinic acid (ALA) on leaf photosynthesis and fruit quality of apples. J. Fruit Sci. 2013, 30, 944–951. [Google Scholar]
- Xie, L.; Cheng, X.H.; Feng, X.X.; Yang, T.; Zhang, Z.P.; Wang, L.J. Effects of an amino acid fertilizer on the leaf photosynthesis and fruit quality of ‘Summer black’ grape. J. Nanjing Agric. Univ. 2013, 36, 31–37. [Google Scholar]
- Wei, J.G.; Yu, H.; Zhang, X.; Jiang, Y.Q.; Zeng, Q.L. Effects of 5-aminolevulinic acid on photosynthetic capacity, yield, and fruit quality of Vaccinium ashei Reade. North. Hortic. 2014, 16, 9–12. [Google Scholar]
- Fang, X.; Zhang, L.Z.; Wang, L.J. The transcription factor MdERF78 is involved in ALA-induced anthocyanin accumulation in apples. Front. Plant Sci. 2022, 13, 915197. [Google Scholar] [CrossRef]
- Liang, R.L.; Wang, L.J.; Wang, X.Q.; Zhang, J.T.; Gan, X. Effects of exogenous ALA on leaf photosynthesis, photosynthate transport, and sugar accumulation in Prunus persica L. Forests 2023, 14, 723. [Google Scholar] [CrossRef]
- Liu, H.; Kang, L.; Wang, L.J. Promotion of 5-aminolevulinic acid on seed germination of watermelon (Citrullus lanatus) under salt stress. J. Fruit Sci. 2006, 1, 854–859. [Google Scholar]
- Wang, L.J.; Jiang, W.B.; Huang, B.J. Promotion of photosynthesis by 5-aminolevulinic acid (ALA) during and after chilling stress in melon seedlings grown under low light condition. Acta Hortic. Sin. 2004, 3, 321–326. [Google Scholar]
- Yin, L.L.; Yu, X.C.; Wang, Y.H.; Xu, Z.H.; Li, K.; Han, D.J. Effect of 5-aminolevulinic acid on chilling tolerance in cucumber seedlings. Acta Agric. Boreali-Occident. Sin. 2007, 16, 166–169. [Google Scholar]
- Liu, T.; Xua, J.J.; Li, J.M.; Hu, X.H. NO is involved in JA- and H2O2-mediated ALA-induced oxidative stress tolerance at low temperatures in tomato. Environ. Exp. Bot. 2019, 161, 334–343. [Google Scholar] [CrossRef]
- Ma, N.; Qi, L.; Gao, J.J.; Chao, K.C.; Hu, Q.F.; Jiang, H.G.; Wang, L.G. Effects of 5-aminolevulinic acid on cutting growth under high temperature condition and leaf chlorophyll fast fluorescence characteristics of Ficus carica L. J. Nanjing Agric. Univ. 2015, 38, 546–553. [Google Scholar]
- An, Y.Y.; Zhang, L.Y.; Feng, X.X.; Tian, F.; Li, J.; Wang, L.J. Effects of 5-aminolevulinic acid on low light tolerance of apple leaves. Acta Bot. Boreal-Occident. Sin. 2016, 36, 987–995. [Google Scholar]
- Long, S.; Liu, B.; Gong, J.; Wang, R.; Gao, S.; Zhu, T.; Guo, H.; Liu, T.; Xu, Y. 5-Aminolevulinic acid promotes low-light tolerance by regulating chloroplast ultrastructure, photosynthesis, and antioxidant capacity in tall fescue. Plant Physiol. Biochem. 2022, 190, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, T.; Gao, W.R.; Guo, S.R.; Li, D.C.; Sun, Y.J. Effects of 5-aminolevulinic (ALA) on growth and photosynthesis of pepper under low temperature stress. Jiangsu J. Agric. Sci. 2011, 27, 612–616. [Google Scholar]
- Wu, Y.; Jin, X.; Liao, W.B.; Hu, L.L.; Dawuda, M.M.; Zhao, X.J.; Tang, Z.Q.; Gong, T.Y.; Yu, J.H. 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.P.; Wang, L.J. Effects of 5-aminolevulinic (ALA) on chlorophyll fluorescence dynamics of watermelon seedlings under shade condition. Acta Hortic. Sin. 2007, 34, 901–908. [Google Scholar]
- Zhao, Y.Y.; Yan, F.; Hu, L.P.; Zhou, X.T.; Zou, Z.R. Effects of 5-aminolevulinic acid on photosynthetic characteristics of tomato seedlings under NaCl stress. Chin. J. Appl. Ecol. 2014, 25, 2919–2926. [Google Scholar]
- Wang, Y.X.; Chen, X.Y.; Li, X.Y.; Song, Y.H.; Wang, J.A.; Huang, J.; Xue, R.L. Exogenous application of 5-aminolevulinic acid alleviated damage to wheat chloroplast ultrastructure under drought stress by transcriptionally regulating genes correlated with photosynthesis and chlorophyll biosynthesis. Acta Physiol. Plant. 2022, 44, 12. [Google Scholar] [CrossRef]
- Chen, Z.; An, Y.; Wang, L. ALA reverses ABA-induced stomatal closure by modulating PP2AC and SnRK2.6 activity in apple leaves. Hortic. Res. 2023, 10, 37287446. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, J.T.; Wang, L.J. ALA induces stomatal opening through regulation among PTPA, PP2AC, and SnRK2.6. Front. Plant Sci. 2023, 14, 1206728. [Google Scholar] [CrossRef]
- Shen, M.; Duan, C.H.; Zhang, Y.P.; Cheng, Y.; Wang, L.J.; Li, B.J. Effects of exogenous ALA treatment on thinning and fruit quality in ‘Hosui’ pear (Pyrus pyrifolia). Acta Hortic. Sin. 2011, 38, 1515–1522. [Google Scholar]
- Shen, M.; Chen, X.H.; Xie, L.; Li, C.; Sun, X.E.; Zhang, Z.P.; Wang, L.J. Effects of amino-acid fertilizer on photosynthesis in leaves of pear cultivars. J. Nanjing Agric. Univ. 2012, 35, 81–86. [Google Scholar]
- Shen, M.; Zhang, Z.P.; Wang, L.J. Effect of 5-aminolevulinic acid (ALA) on leaf diurnal photosynthetic characteristics and antioxidant activity in pear (Pyrus pyrifolia Nakai). In Artificial Photosynthesis; Najafpour, M.M., Ed.; InTech: London, UK, 2011; pp. 239–265. [Google Scholar]
- Wang, P.F.; Zhao, J.X.; Li, T.; Yu, L.C.; Li, Y.F.; Jia, Y.L.; Zhang, Y.X.; Zhang, H.X.; Quan, C.; Xu, J.F.; et al. Study on the effect of exogenous ALA to alleviate low temperature injury in pear flower organs. China Fruits 2023, 1, 16–21. [Google Scholar]
- Xiao, C.Z.; Zhang, S.L.; Hu, H.J.; Tian, R.; Wu, J.; Yang, Z.J.; Ma, C.Y. Effects of bagging and exogenous 5-aminolevulinic acid treatment on coloration of ‘Yunhongli 2’. J. Nanjing Agric. Univ. 2012, 35, 25–29. [Google Scholar]
- Li, W.X.; Cao, X.Y.; Wang, X.Q. Effect of ALA combined with calcium chloride and sucrose on fruit quality and color of ‘Nanhong’ pear. China Fruits 2022, 1, 35–38. [Google Scholar]
- T/JAASS 88-2023; 5-Aminolevulic Acid Technical Procedures for Application in Peach. Jiangsu Association of Agricultural Science Societies: Nanjing, China, 2023.
- Wang, X.K. Experimental Principles and Techniques of Plant Physiology and Biochemistry; Higher Education Press: Beijing, China, 2006; pp. 130–134. [Google Scholar]
- Harel, E.; Klein, S. Light dependent formation of δ-aminolevulinic acid in etiolated of higher plants. Biochem. Biophys. Res. Commun. 1972, 49, 364–370. [Google Scholar] [CrossRef]
- Li, P.M.; Gao, H.Y. Application of rapid chlorophyll fluorescence induction kinetics analysis in the study of photosynthesis. J. Plant Physiol. Mol. Biol. 2005, 1, 559–566. [Google Scholar]
- Wang, D.Y.; Li, X.; Xu, L.; An, Y.Y.; Wang, L.J. 5-Aminolevulinic acid (ALA) improved the heat resistance of leaves of two garden plants. J. Bot. Res. 2018, 73, 350–365. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, Y.J.; Jiang, B.; Zhang, Y.; Li, Y.B.; He, L.; Wang, Y.C. Transient expression in tobacco by Agrobacterium mediated transformmation. J. Northeast. For. Univ. 2010, 38, 110–112. [Google Scholar]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Hotta, Y.; Tanaka, H.; Takaoka, Y.; Takeuchi, Y.; Konnai, M. Promotive effects of 5-aminolevulinic acid on the yield of several crops. Plant Growth Regul. 1997, 22, 109–114. [Google Scholar] [CrossRef]
- Liu, W.Q.; Kang, L.; Wang, L.J. Effects on strawberry photosynthesis and relations to anti-oxidant enzymes of ALA. Acta Bot. Boreali-Occident. Sin. 2006, 1, 57–62. [Google Scholar]
- Wang, J.N.; Li, X.Y.; Wei, S.M.; Zhao, H.J.; Zhao, M.Q.; Wang, Y.X. Regulation of exogenous 5-aminolevulinic acid on photosynthesis and D1 protein of wheat seedlings under drought stress. Crop J. 2018, 1, 121–126. [Google Scholar]
- Wang, L.J.; Zhang, J.T.; Zhong, Y.; Zhang, L.Z.; Yang, H.; Liu, L.B.; Zhou, J.Y.; Iqbal, M.M.; Gan, X. Regulation of 5-aminolevunilic acid and its application in agroforestry. Forests 2023, 14, 1857. [Google Scholar] [CrossRef]
- Chen, L.H.; Liu, L.B.; An, Y.Y.; Zhang, Z.P.; Wang, L.J. Preliminary studies on the possible mechanism underlying 5-aminolevulinic acid-induced stomatal opening in apple leaves. Acta Hortic. Sin. 2014, 41, 1965–1974. [Google Scholar]
- Liu, L.B.; An, Y.Y.; Xiong, L.J.; Wang, L.J. Flavonols induced by 5-aminolevulinic acid are involved in regulation of stomatal opening in apple leaves. Acta Hortic. Sin. 2016, 43, 817–828. [Google Scholar] [CrossRef]
- Xiong, L.J.; An, Y.Y.; Wang, L.J. The role of microtubule skeleton and PP1/PP2A protein phosphatase in ALA-ABA regulating stomatal movement in apple leaves. Acta Hortic. Sin. 2018, 45, 2073–2088. [Google Scholar]
- Yang, Y.X.; Xia, J.X.; Fang, X.; Jia, H.R.; Wang, X.C.; Lin, Y.L.; Liu, S.Y.; Ge, M.Q.; Pu, Y.F.; Fang, J.G.; et al. Drought stress in ‘Shine Muscat’ grapevine: Consequences and a novel mitigation strategy-5-aminolevulinic acid. Front. Plant Sci. 2023, 14, 1129114. [Google Scholar] [CrossRef]
- Fromme, P.; Melkozernov, A.; Jordan, P.; Krauss, N. Structure and function of photosystem I: Interaction with its soluble electron carriers and external antenna systems. FEBS Lett. 2003, 555, 40–44. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tanaka, A.; Tsuji, H. Effects of 5-aminolevulinic acid on the accumulation of chlorophyll-b and apoproteins of the light-harvesting chlorophyll a/b-protein complex of photosystem II. Plant Cell Physiol. 1993, 34, 465–472. [Google Scholar]
- Wang, L.J.; Jiang, W.B.; Zhang, Z.; Yao, Q.H.; Matsui, H.; Ohara, H. Biosynthesis and physiological activity of 5-aminolevulinic acid and its potential application in agriculture. Plant Physiol. Commun. 2003, 3, 185–192. [Google Scholar]
- Zhong, Y.; Liu, C.; Wei, B.; Zhang, J.; An, Y.; Wang, L. Exogenous 5-aminolevulinic acid promotes osmotic stress tolerance of walnuts by modulating photosynthesis, osmotic adjustment, and antioxidant systems. Forests 2023, 14, 789. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Li, J.; Dawuda, M.M.; Ali, B.; Wu, Y.; Yu, J.; Tang, Z.; Lyu, J.; Xiao, X.; et al. Exogenous application of 5-aminolevulinic acid promotes coloration and improves the quality of tomato fruit by regulating carotenoid metabolism. Front. Plant Sci. 2021, 12, 683868. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wang, J.W.; Li, R.R.; Tian, Q.; Wu, Y.; You, J.H. The mitigation effect of exogenous ALA on salt stress of tomato seedlings. China Cucurbits Veg. 2023, 36, 56–64. [Google Scholar]
- Wang, H.; Liu, Z.; Luo, S.; Li, J.; Zhang, J.; Li, L.; Xie, J. 5-Aminolevulinic acid and hydrogen sulphide alleviate chilling stress in pepper (Capsicum annuum L.) seedlings by enhancing chlorophyll synthesis pathway. Plant Physiol. Biochem. 2021, 167, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.R.; Zhang, F.T.; Gao, J.X.; Sun, X.Q.; Deng, X.J. An overview of chlorophyll biosynthesis in higher plants. Acta Bot. Boreali-Occident. Sin. 2009, 29, 629–636. [Google Scholar]
- Wang, R.J.; Jiang, Y.; Zhu, F.; Tang, Z.; Liu, X.S.; Jiang, Y.; Wang, Y. Application of 820 nm optical absorption curve technology to study the performance of leaf optical system I under stress conditions. Guangxi For. Sci. 2020, 49, 152–155. [Google Scholar]
- Sun, Y.P.; Liu, J.; Cao, X.; Huang, Y.J.; Hall, A.M.; Guo, C.B.; Wang, L.J. Effects of 5-aminolevulinic acid treatment on photosynthesis of strawberry. Photosynthetica 2017, 55, 276–284. [Google Scholar] [CrossRef]
- Song, Q.N.; Feng, P.W.; Liu, Y.; Yang, X.H. The research progress of the mechanism on PSII assemble and repair circulation. Plant Physiol. J. 2019, 55, 133–140. [Google Scholar]
- Hou, P.F.; Ma, J.Q.; Zhao, P.F.; Zhang, H.L.; Zhao, H.J.; Liu, H.S.; Zhao, Y.D.; Wang, Y.X. Effects of betaine on chloroplast protective enzymes and psbA gene expression in wheat seedlings under drought stress. Acta Agron. Sin. 2013, 39, 1319–1324. [Google Scholar] [CrossRef]
- Wang, M.; Shan, J.X.; Zhong, Z.P.; Li, L.B.; Kang, T.Y. Advances in the research of structure and function of photosystem ii core antenna complexes CP43 and CP47. Chin. Bull. Bot. 2000, 17, 141–149. [Google Scholar]
- Jia, H.S.; Li, D.Q.; Ham, Y.Q. Advances in studies on photoinhibition in photosynthesis of higher plants. Chin. Bull. Bot. 2000, 17, 218–224. [Google Scholar]
Figure 1.
Effect of ALA treatment on photosynthetic gas exchange parameters of pear leaves. (A) Pn: net photosynthetic rate. (B) Tr: transpiration rate. (C) Gsw: stomatal conductance to water vapor. (D) Ci: intercellular CO2 concentration. (E) Pn/Ci: instantaneous carboxylation efficiency. (F) WUE: water use efficiency. The data are the means of twenty biological repeats ± SE. The same lowercases in each panel represent no significant difference at p = 0.05.
Figure 1.
Effect of ALA treatment on photosynthetic gas exchange parameters of pear leaves. (A) Pn: net photosynthetic rate. (B) Tr: transpiration rate. (C) Gsw: stomatal conductance to water vapor. (D) Ci: intercellular CO2 concentration. (E) Pn/Ci: instantaneous carboxylation efficiency. (F) WUE: water use efficiency. The data are the means of twenty biological repeats ± SE. The same lowercases in each panel represent no significant difference at p = 0.05.

Figure 2.
Effect of ALA treatment on the chlorophyll fast fluorescence curves (A–E) and the 820 nm modulated reflection fluorescence curves (F–J) of pear leaves. The capitals O, J, I, and P in the A represent the fluorescence at 0 ms, 3 ms, 30 ms, and 300 ms, respectively. The data are the means of fifteen biological repeats.
Figure 2.
Effect of ALA treatment on the chlorophyll fast fluorescence curves (A–E) and the 820 nm modulated reflection fluorescence curves (F–J) of pear leaves. The capitals O, J, I, and P in the A represent the fluorescence at 0 ms, 3 ms, 30 ms, and 300 ms, respectively. The data are the means of fifteen biological repeats.

Figure 3.
Effect of exogenous ALA spraying on the endogenous ALA content (A) and the biosynthetic (B) and catabolic capacity (C) of pear leaves. The data are the means of three biological repeats ± SE. The same lowercases in each panel represent no significant difference at p = 0.05.
Figure 3.
Effect of exogenous ALA spraying on the endogenous ALA content (A) and the biosynthetic (B) and catabolic capacity (C) of pear leaves. The data are the means of three biological repeats ± SE. The same lowercases in each panel represent no significant difference at p = 0.05.

Figure 4.
Effect of ALA treatment on the content of photosynthetic pigment in pear leaves. (A) Relative content of chlorophylls (SPAD), (B) chlorophyll a, (C) chlorophyll b, (D) total chlorophyll content, (E) ration of Chl a/b, (F) carotenoid content. The data are the means of nine biological repeats ± SE. The same lowercases in each panel represent no significant difference at p = 0.05.
Figure 4.
Effect of ALA treatment on the content of photosynthetic pigment in pear leaves. (A) Relative content of chlorophylls (SPAD), (B) chlorophyll a, (C) chlorophyll b, (D) total chlorophyll content, (E) ration of Chl a/b, (F) carotenoid content. The data are the means of nine biological repeats ± SE. The same lowercases in each panel represent no significant difference at p = 0.05.

Figure 5.
Chlorophyll biosynthesis pathway and the effect of ALA treatment on the expression of related genes in pear leaves. C1 to C5 and A1 to A5 represent the control and ALA treatment collected from 1 to 5 weeks after ALA foliar spraying, respectively. The color gradient from blue to red corresponds to fold changes from low to high. Glu: glutamic acid; ALA: 5-aminolevulinic acid; PBG: porphobilinogen; HMB: hydroxymethylbilane; Urogen III: uroporphyrnogen III; coprogen III: coproporphyrnogen III; Proto IX: protoporphyrnogen IX; MgP IX: mg-protoporphyrin IX; MgPME: Mg-protoporphyrin IX methyl ester; Pchlide: protochlorophyllide; Chlide: chlorophyllide; Chl a: chlorophyllide a; Chl b: chlorophyllide b. The genes depicted in the heatmap correspond to enzymes marked with green highlights in the chlorophyll biosynthesis pathway.
Figure 5.
Chlorophyll biosynthesis pathway and the effect of ALA treatment on the expression of related genes in pear leaves. C1 to C5 and A1 to A5 represent the control and ALA treatment collected from 1 to 5 weeks after ALA foliar spraying, respectively. The color gradient from blue to red corresponds to fold changes from low to high. Glu: glutamic acid; ALA: 5-aminolevulinic acid; PBG: porphobilinogen; HMB: hydroxymethylbilane; Urogen III: uroporphyrnogen III; coprogen III: coproporphyrnogen III; Proto IX: protoporphyrnogen IX; MgP IX: mg-protoporphyrin IX; MgPME: Mg-protoporphyrin IX methyl ester; Pchlide: protochlorophyllide; Chlide: chlorophyllide; Chl a: chlorophyllide a; Chl b: chlorophyllide b. The genes depicted in the heatmap correspond to enzymes marked with green highlights in the chlorophyll biosynthesis pathway.

Figure 6.
Effect of exogenous ALA on the promoter activities of genes associated with ALA and chlorophyll synthesis. (A) Construction of gene vector atlas for GUS staining analysis. (B) Analysis of GUS gene expression in tobacco leaves after injection with different agrobacterium liquids. (C) Analysis of GUS expression in tobacco leaves after injection of different Agrobacterium solutions. Empty represents the GUS expressions of tobacco injected with 35S::GUS Agrobacterium solution, while the control and ALA respectively represent the GUS expressions in the tobacco injected with Agrobacterium liquids with or without ALA. Different lowercase letters in each histogram represent significant differences at p = 0.05.
Figure 6.
Effect of exogenous ALA on the promoter activities of genes associated with ALA and chlorophyll synthesis. (A) Construction of gene vector atlas for GUS staining analysis. (B) Analysis of GUS gene expression in tobacco leaves after injection with different agrobacterium liquids. (C) Analysis of GUS expression in tobacco leaves after injection of different Agrobacterium solutions. Empty represents the GUS expressions of tobacco injected with 35S::GUS Agrobacterium solution, while the control and ALA respectively represent the GUS expressions in the tobacco injected with Agrobacterium liquids with or without ALA. Different lowercase letters in each histogram represent significant differences at p = 0.05.

Figure 7.
PSII reaction center structure and the effect of ALA treatment on the expressions of related genes in pear leaves. C1 to C5 and A1 to A5 represent the samples collected 1 to 5 weeks from the control and ALA foliar spraying for 1 to 5 weeks, respectively. The color gradient from blue to red corresponds to fold changes from low to high.
Figure 7.
PSII reaction center structure and the effect of ALA treatment on the expressions of related genes in pear leaves. C1 to C5 and A1 to A5 represent the samples collected 1 to 5 weeks from the control and ALA foliar spraying for 1 to 5 weeks, respectively. The color gradient from blue to red corresponds to fold changes from low to high.

Figure 8.
Effect of exogenous ALA on the promoter activities of genes associated with PSII. (A) Construction of gene vector atlas for GUS qualitative and quantitative analysis. (B) Analysis of GUS gene expression in tobacco leaves after injection with different agrobacterium liquids. (C) Analysis of GUS expression in tobacco leaves after injection of different Agrobacterium solutions. Empty represents the GUS expressions of tobacco injected with 35S::GUS Agrobacterium solution, while ALA and control represent the GUS expressions in the tobacco injected with Agrobacterium liquids treated with or without ALA, respectively. Different lowercase letters in each histogram represent significant differences at p = 0.05.
Figure 8.
Effect of exogenous ALA on the promoter activities of genes associated with PSII. (A) Construction of gene vector atlas for GUS qualitative and quantitative analysis. (B) Analysis of GUS gene expression in tobacco leaves after injection with different agrobacterium liquids. (C) Analysis of GUS expression in tobacco leaves after injection of different Agrobacterium solutions. Empty represents the GUS expressions of tobacco injected with 35S::GUS Agrobacterium solution, while ALA and control represent the GUS expressions in the tobacco injected with Agrobacterium liquids treated with or without ALA, respectively. Different lowercase letters in each histogram represent significant differences at p = 0.05.


{kind=link}
{kind=link}
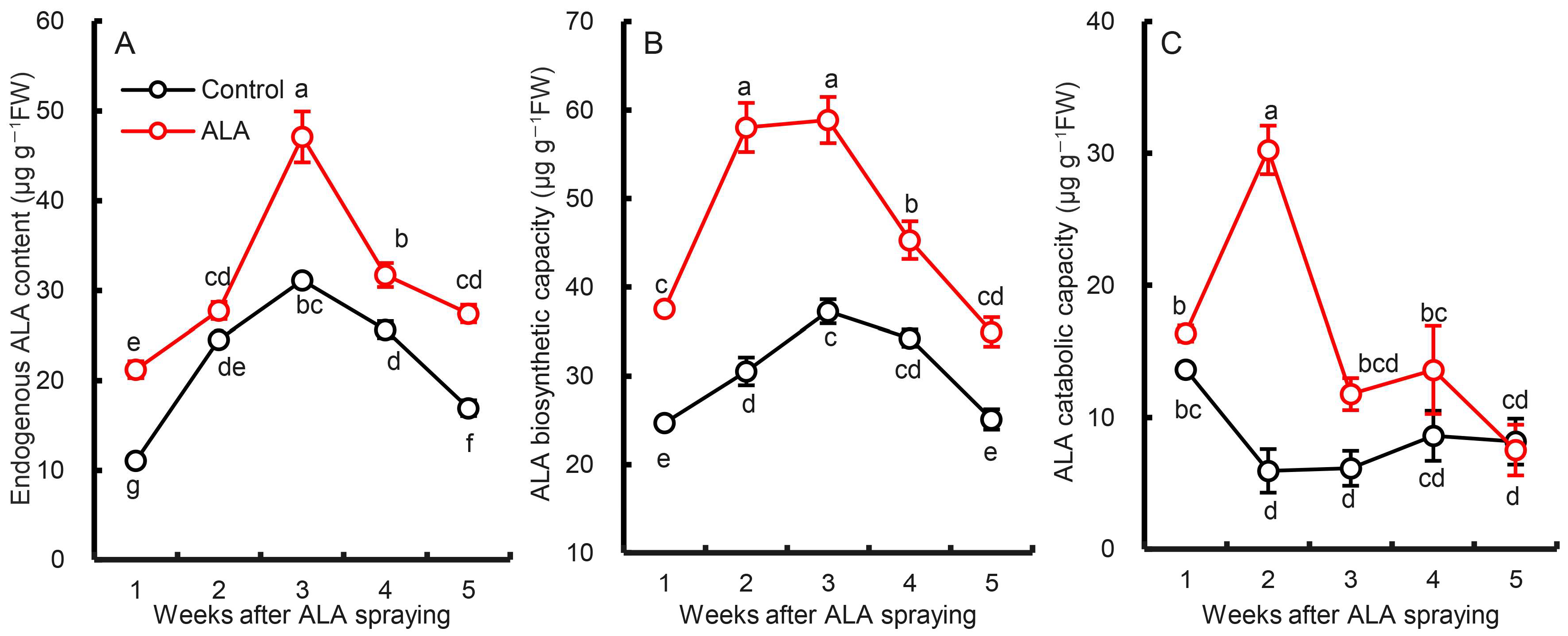{kind=link}
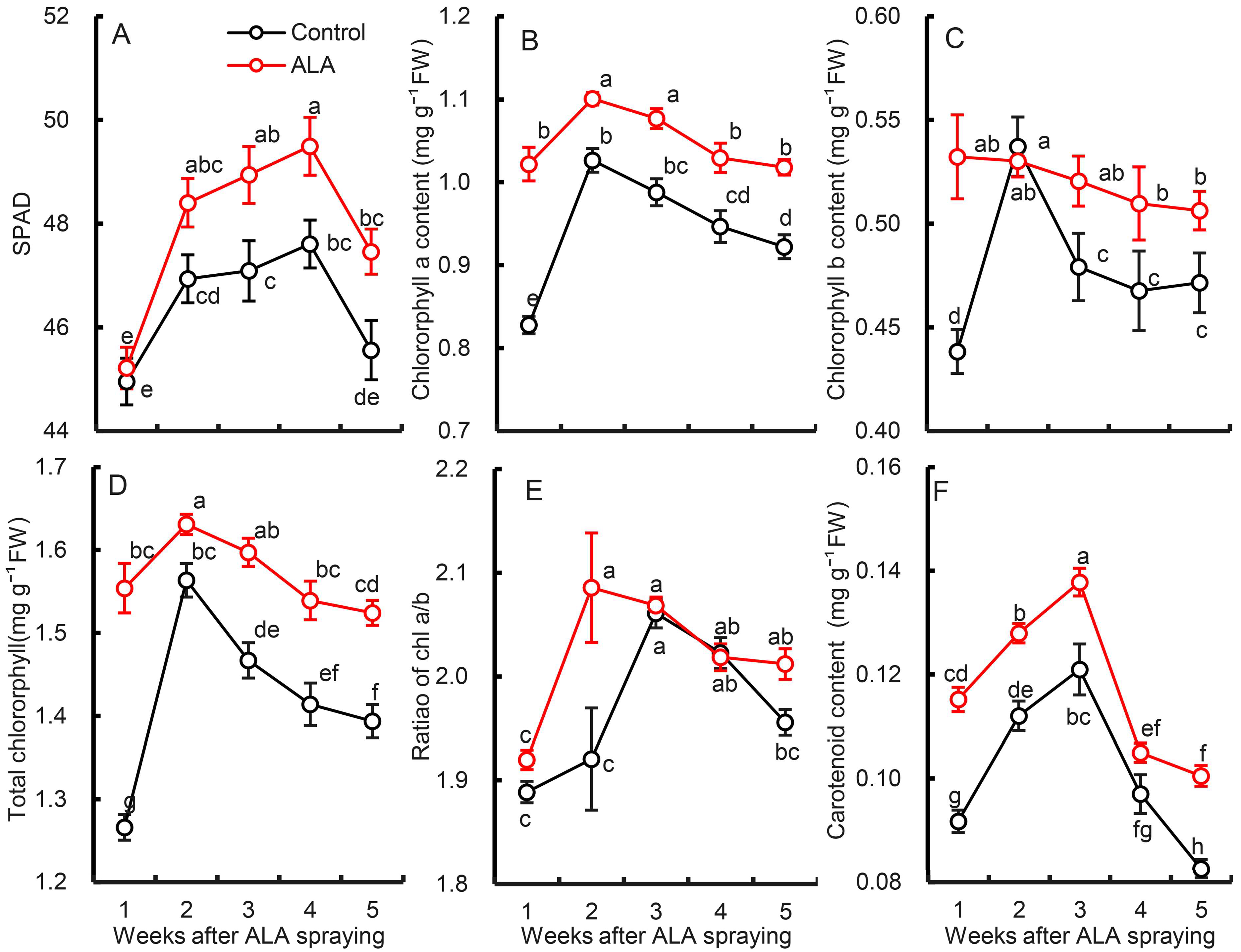{kind=link}
{kind=link}
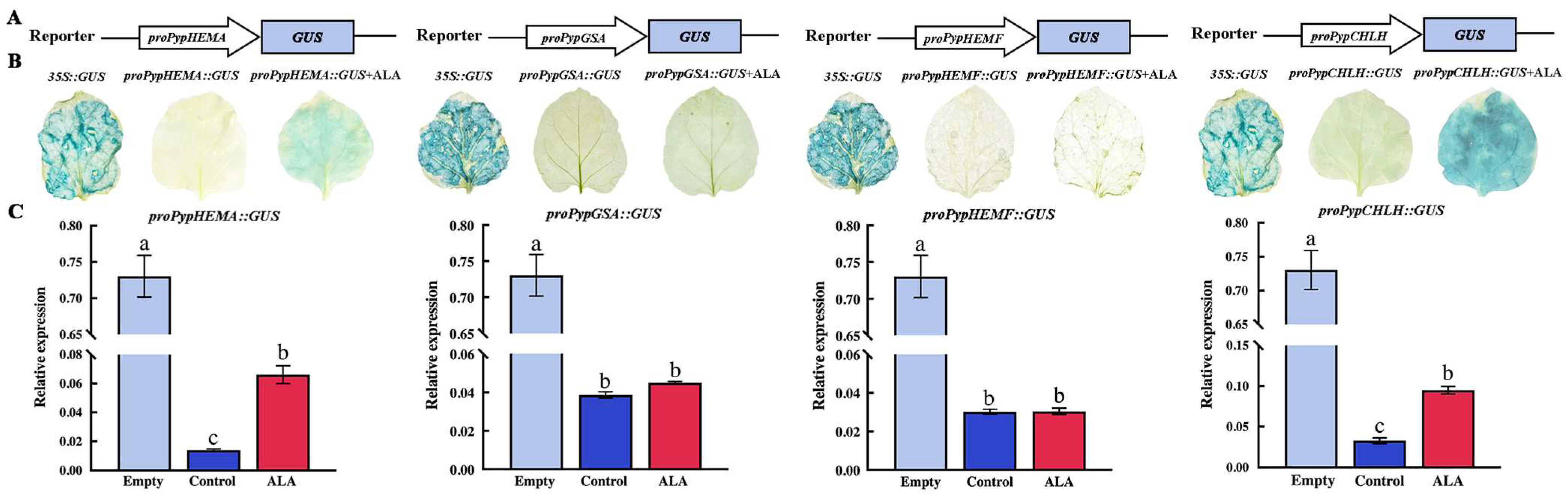{kind=link}
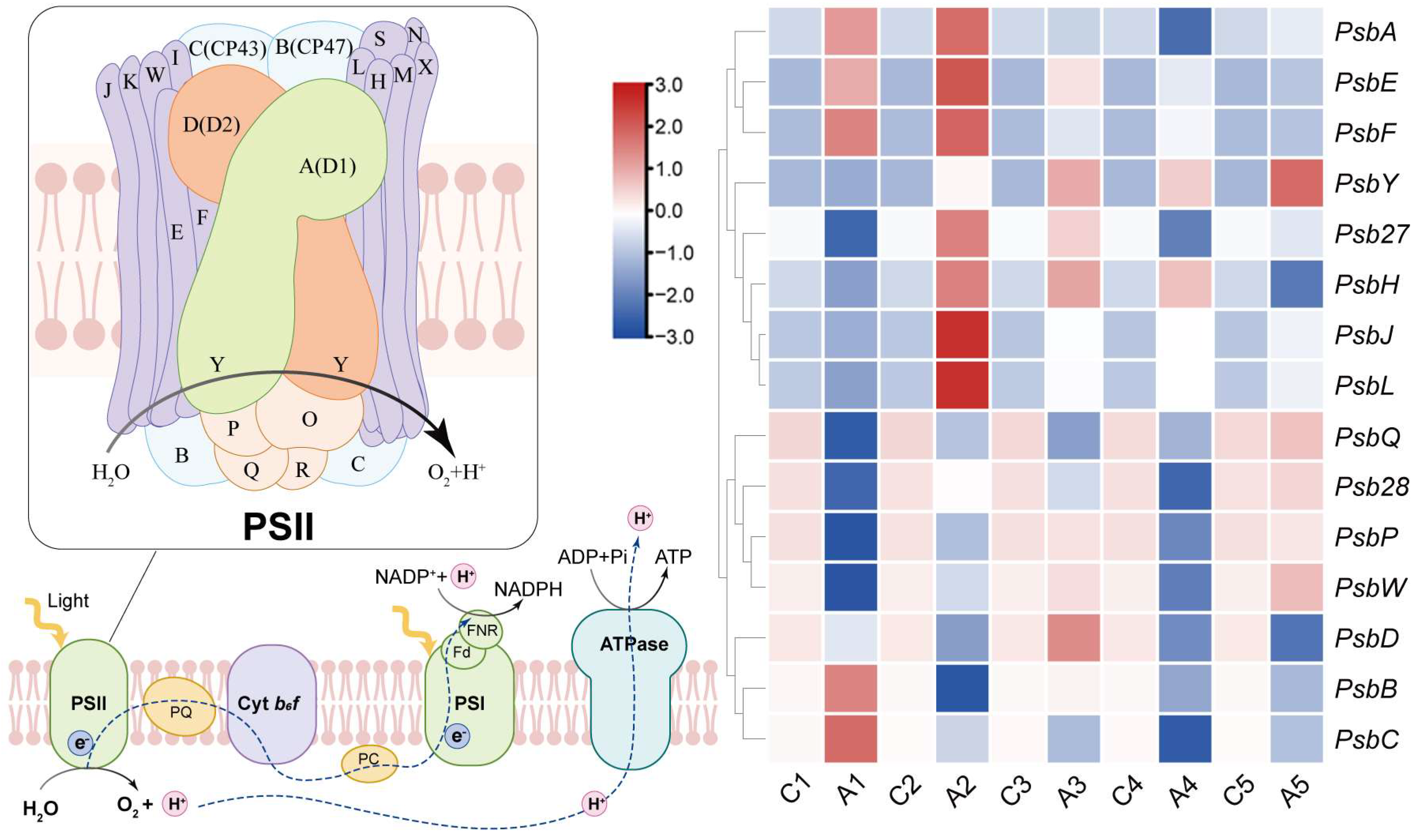{kind=link}
{kind=link}
Table 1.
Primer sequences used in this experiment.
| Name (Gene ID) | Primer | Amplicon Size |
|---|---|---|
| Actin (LOC103931062) | Fw:cccagaagtgctcttccaac Rv:ttgatcttcatgctgcttgg | 1550 |
| PsbA (ON478189.1:308-1369) | Fw:cattttctgtggtttccctgat Rv:cgcccttggattgctgtt | 1061 |
| PsbB (ON478189.1:75738-77264) | Fw:tttcgggcttgtgcttttt Rv:cccacgctggatttacgg | 1526 |
| PsbC (ON478189.1:36669-38088) | Fw:ggcgggggagatgtaagaa Rv:ccaaacaaagcaacaagcaata | 1419 |
| PsbD (ON478189.1:35660-36721) | Fw:atgggagttgctggggtg Rv:ccaaaaacggttagcggtg | 1061 |
| PsbE (ON478189.1:67489-67740) | Fw:gaattccttgtcggctttctg Rv:ccttatttattgcgggttggt | 251 |
| PsbF (ON478189.1:c67479-67360) | Fw:gtacctaccgtttcttttttaggg Rv:ttggatgaactgcattgctga | 119 |
| PsbH (ON478189.1:77867-78091) | Fw:gatctggccgcccaaaa Rv:tgcgagacccatcaaagga | 224 |
| PsbJ (ON478189.1:c67094-66972) | Fw:acgacacaatcaaacccgaa Rv:caaaaatgagtaataacccccag | 122 |
| PsbL (ON478189.1:c67337-67221) | Fw:acgacacaatcaaacccgaa Rv:caaaaatgagtaataacccccag | 116 |
| PsbP (LOC103965639) | Fw:ctgagtccgagggtggctt Rv:tccttcatctccgtctgctgt | 807 |
| PsbQ (LOC103933887) | Fw:cttgctgtctgaaacaaccctc Rv:ttctctccacgactcgaactct | 674 |
| PsbW (LOC103946003) | Fw:tggaaggaaaaaaggagagca Rv:cgagggaggaagtatagataaagtaga | 623 |
| PsbY (LOC103949857) | Fw:caaccgcatgacaaacaagag Rv:ccaccacaaacagcaacaact | 887 |
| Psb27 (LOC103947836) | Fw:ccacccccacaatccca Rv:ccttatttatcacctcctccgtt | 874 |
| Psb28 (LOC103959875) | Fw:tcaggcttgttatgttgggg Rv:tctggtattgtctgttcatcggt | 987 |
| HEMA (LOC103957838) | Fw:acaaggaggataggctaaggaag Rv:ccattggaccatgaaggagtt | 2115 |
| GSA (LOC103936176) | Fw:taatggtatccgcaagctcac Rv:aacttcctggctccctcaac | 1716 |
| HEMB (LOC125472965) | Fw:gttgaggcaaatgaagacgagt Rv:tgcagctataggcagaggagag | 1537 |
| HEMD (LOC103957047) | Fw:tgctatgacaggagcggtga Rv:cctaagaatgggattgaaaaatg | 1367 |
| HEMF (LOC125474519) | Fw:caccgccccgatacctt Rv:tgacaccagccttctccca | 1485 |
| HEMG (LOC103967516) | Fw:atgatggtggactgtggtttga Rv:gacctggaggaggcgga | 2010 |
| CHLH (LOC103946503) | Fw:ggaggcaagaggggcg Rv:gggcaaaaccagtaagcga | 4452 |
| CHLM (LOC103931332) | Fw:ccaagaccgttgagaatgtga Rv:gccctgccgagaagcc | 1288 |
| CRD1 (LOC103938880) | Fw:agacaaatcgcactacagcca Rv:ttcaagtttttcaccacggg | 1424 |
| DVR (LOC103958557) | Fw:cgtggcaaaaacccaaaag Rv:aacacccaaatcagtcacatcc | 1531 |
| POR (LOC103949278) | Fw:ccctccgactgcctctca Rv:tactggcttccacttctgcttat | 1843 |
| CHLG (LOC103965133) | Fw:ttgaaggagatagagcaatggg Rv:tggaaaaagacttggggagc | 1470 |
| CAO (LOC103940306) | Fw:cccttctcccagcacttctac Rv:gacacatccaggtttcccatc | 2157 |
| NtActin (LOC107830629) | Fw:cctgaggtccttttccaacca Rv:ggattccggcagcttccatt | 1574 |
| GUS (LOC103930738) | Fw:cctgaggtccttttccaacca Rv:tcattgtttgcctccctgct | 2038 |
| Promoter of PsbA | Fw:accatgattacgccaagcttgatatccaccatcttgacttggtgatgg Rv:accacccggggatcctctagaatggagagagagagatattggagttgaagg | 254 |
| Promoter of PsbB | Fw:accatgattacgccaagcttctaattgctacacgcattgcagaatg Rv:accacccggggatcctctagatgccaatcagcttctctccacc | 502 |
| Promoter of PsbJ | Fw:accatgattacgccaagcttcgtaccaataacgtccaaatgattggtaca Rv:accacccggggatcctctagaaccaggactttagacaacggattacgtag | 1032 |
| Promoter of HEMA | Fw:accatgattacgccaagcttggacttttttttttttgggagaggagct Rv:accacccggggatcctctagaagggagggatatggaattgggataagg | 2000 |
| Promoter of GSA | Fw:accatgattacgccaagcttccccctgtagtattacaaattaaaaatttt Rv:accacccggggatcctctagaagactttggtcttagtatatatgagttctt | 2000 |
| Promoter of HEMF | Fw:accatgattacgccaagcttgtactatatctcactttgatgtgaaagcg Rv:accacccggggatcctctagatggaaatacccctttatcaatgaaaaatag | 2000 |
| Promoter of CHLH | Fw:accatgattacgccaagcttgatctgcggatttatcaacatcaagctc Rv:accacccggggatcctctagaataatcggatgaactgagttgtagacatga | 2000 |
Table 2.
Effects of ALA treatment on chlorophyll fluorescence parameters of pear leaves.
| Weeks after Foliar Spraying of ALA | Treatment | Wk | Mo | Sm | φo | φPo | φEo | φDo | RC/CS (×103) | PIABS | PItotal | VPSI (×10−4) | VPSII-PSI (×10−5) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Control | 0.37 ± 0.01 ab | 0.53 ± 0.01 a | 28.21 ± 0.74 b | 0.63 ± 0.01 f | 0.80 ± 0.01 d | 0.54 ± 0.00 f | 0.16 ± 0.00 a | 4.48 ± 0.06 bc | 5.77 ± 0.15 c | 4.37 ± 0.23 c | 20.27 ± 0.19 a | 3.52 ± 0.10 abc |
| ALA | 0.35 ± 0.00 d | 0.49 ± 0.01 bc | 30.71 ± 0.69 a | 0.65 ± 0.01 de | 0.85 ± 0.00 b | 0.56 ± 0.01 de | 0.15 ± 0.00 bc | 4.73 ± 0.03 a | 6.18 ± 0.13 abc | 5.04 ± 0.17 ab | 19.90 ± 0.31 ab | 3.86 ± 0.16 a | |
| 2 | Control | 0.37 ± 0.00 b | 0.50 ± 0.01 b | 27.25 ± 0.64 b | 0.66 ± 0.01 cd | 0.80 ± 0.01 d | 0.56 ± 0.00 cd | 0.15 ± 0.00 bc | 4.35 ± 0.03 c | 5.90 ± 0.22 bc | 4.53 ± 0.13 c | 19.13 ± 0.33 bc | 3.11 ± 0.70 d |
| ALA | 0.35 ± 0.00 cd | 0.46 ± 0.01 cd | 30.81 ± 0.71 a | 0.69 ± 0.01 a | 0.85 ± 0.00 ab | 0.59 ± 0.00 a | 0.14 ± 0.00 d | 4.54 ± 0.05 b | 6.61 ± 0.21 a | 5.22 ± 0.13 a | 19.85 ± 0.26 ab | 3.57 ± 0.14 ab | |
| 3 | Control | 0.35 ± 0.01 d | 0.49 ± 0.01 bc | 28.23 ± 0.64 b | 0.67 ± 0.01 bc | 0.81 ± 0.01 d | 0.58 ± 0.00 bc | 0.14 ± 0.00 d | 4.49 ± 0.02 bc | 5.75 ± 0.17 c | 4.67 ± 0.12 bc | 18.17 ± 0.37 de | 3.18 ± 0.19 bcd |
| ALA | 0.33 ± 0.00 e | 0.45 ± 0.01 d | 30.26 ± 0.42 a | 0.69 ± 0.01 a | 0.86 ± 0.00 a | 0.59 ± 0.00 a | 0.14 ± 0.00 e | 4.70 ± 0.07 a | 6.49 ± 0.18 a | 5.25 ± 0.15 a | 18.34 ± 0.27 cde | 3.45 ± 0.12 bcd | |
| 4 | Control | 0.39 ± 0.00 a | 0.50 ± 0.01 b | 23.70 ± 0.50 c | 0.66 ± 0.01 cd | 0.83 ± 0.00 c | 0.56 ± 0.01 d | 0.16 ± 0.00 b | 4.11 ± 0.05 d | 5.95 ± 0.18 bc | 4.97 ± 0.13 ab | 17.56 ± 0.43 e | 2.04 ± 0.10 e |
| ALA | 0.37 ± 0.00 b | 0.48 ± 0.01 bcd | 24.69 ± 0.60 c | 0.68 ± 0.01 ab | 0.85 ± 0.00 ab | 0.58 ± 0.00 ab | 0.15 ± 0.00 c | 4.35 ± 0.07 c | 6.34 ± 0.18 ab | 5.41 ± 0.09 a | 18.35 ± 0.22 cde | 2.33 ± 0.09 e | |
| 5 | Control | 0.36 ± 0.01 bc | 0.50 ± 0.01 b | 26.70 ± 0.68 b | 0.64 ± 0.01 ef | 0.82 ± 0.01 c | 0.55 ± 0.01 ef | 0.15 ± 0.00 bc | 4.51 ± 0.07 b | 5.75 ± 0.16 c | 4.75 ± 0.08 bc | 18.29 ± 0.24 cde | 3.04 ± 0.15 d |
| ALA | 0.35 ± 0.01 cd | 0.48 ± 0.01 bc | 27.86 ± 0.62 b | 0.66 ± 0.01 cd | 0.85 ± 0.01 ab | 0.56 ± 0.01 bcd | 0.14 ± 0.00 d | 4.69 ± 0.05 a | 6.09 ± 0.17 abc | 5.04 ± 0.13 ab | 18.58 ± 0.25 cd | 3.13 ± 0.13 cd |
Note: The data are the means ± SE of 15 biological replicates. The same lowercases in each column represent no significant difference at p = 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Zhang, J.; Wang, L. Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves. Forests 2024, 15, 769. https://0-doi-org.brum.beds.ac.uk/10.3390/f15050769
AMA Style
Wang X, Zhang J, Wang L. Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves. Forests. 2024; 15(5):769. https://0-doi-org.brum.beds.ac.uk/10.3390/f15050769
Chicago/Turabian StyleWang, Xinqing, Jianting Zhang, and Liangju Wang. 2024. "Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (ALA) in Regulating the Photosynthetic Efficiency of Pear Leaves" Forests 15, no. 5: 769. https://0-doi-org.brum.beds.ac.uk/10.3390/f15050769
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.