
The Influence of Tacrolimus on Cellular Morphology, Cellular Viability, Osteogenic Differentiation, and mRNA Expression within Stem Cell Spheroids

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Spheroids Made from Gingiva-Derived Mesenchymal Stem Cells
2.2. The Assessment of Cellular Viability
2.3. Levels of Alkaline Phosphatase Activity and Calcium Deposition
2.4. Total RNA Extraction and Real-Time Polymerase Chain Reaction Quantification
2.5. Western Blot Analysis
2.6. RNA Isolation, Library Preparation, Sequencing and Data Analysis
2.7. Statistical Analysis
3. Results
3.1. Creation of Stem Cell Aggregates in Spheroid Form
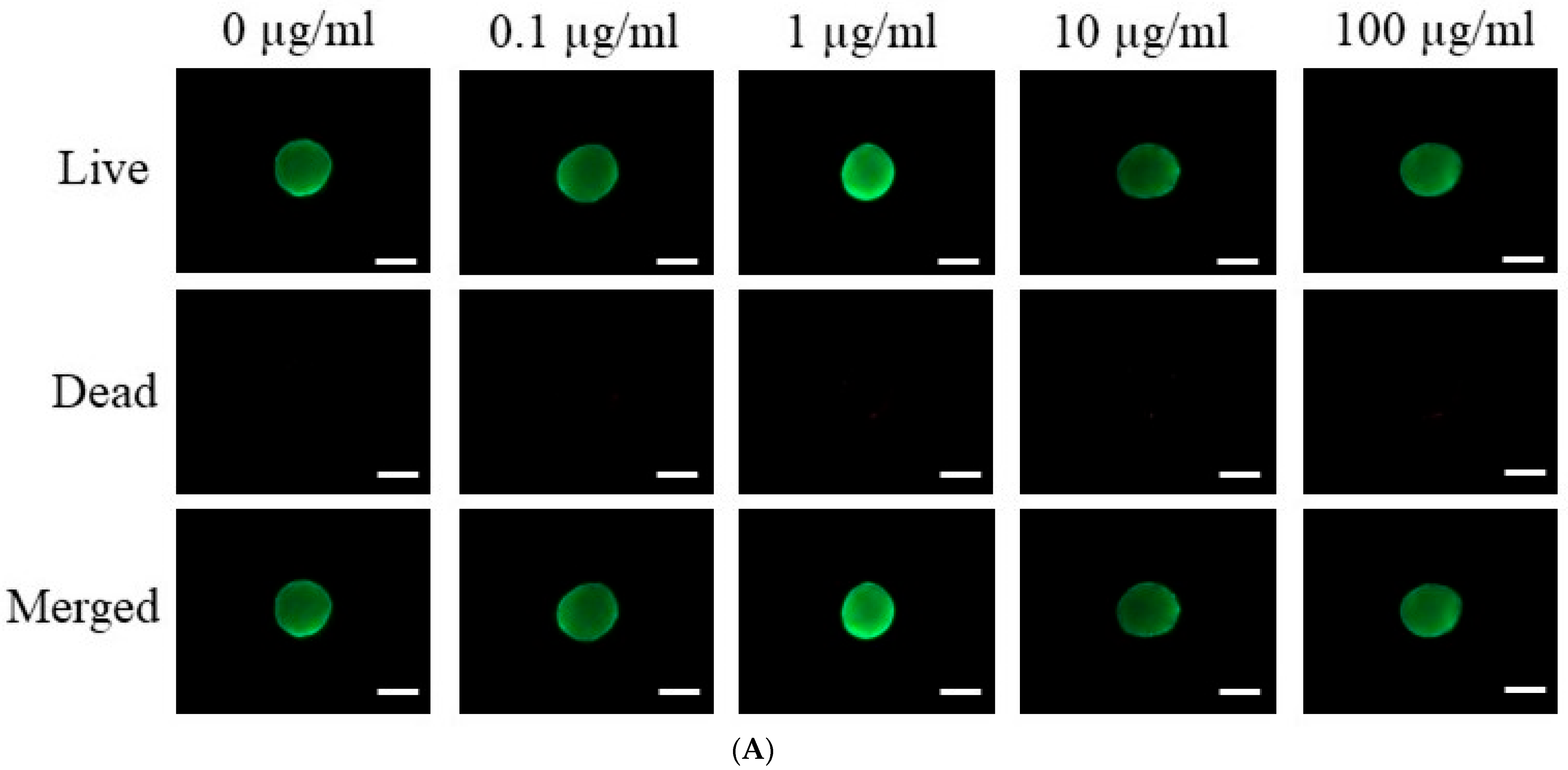
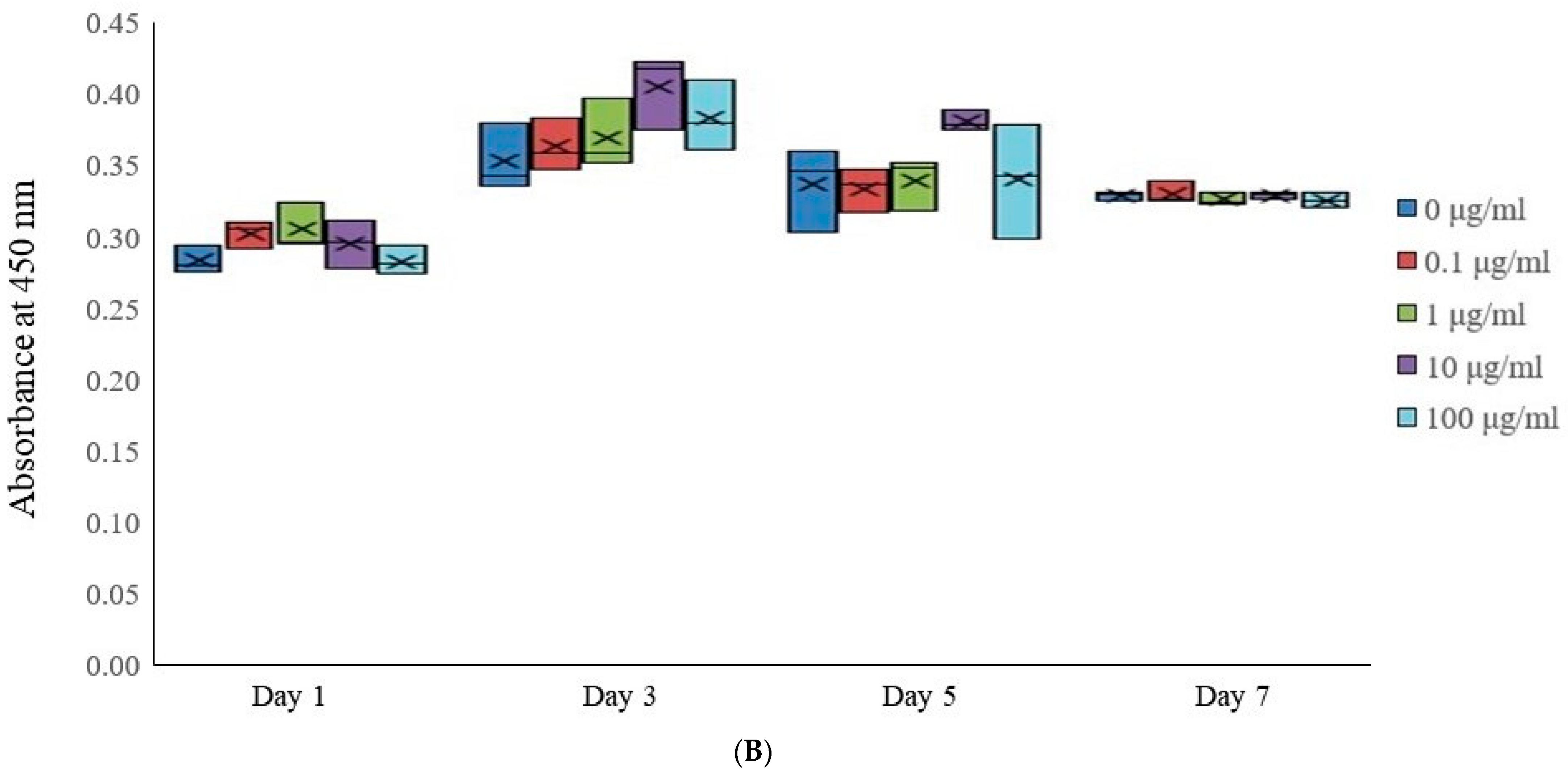
3.2. Assessment of Cell Viability
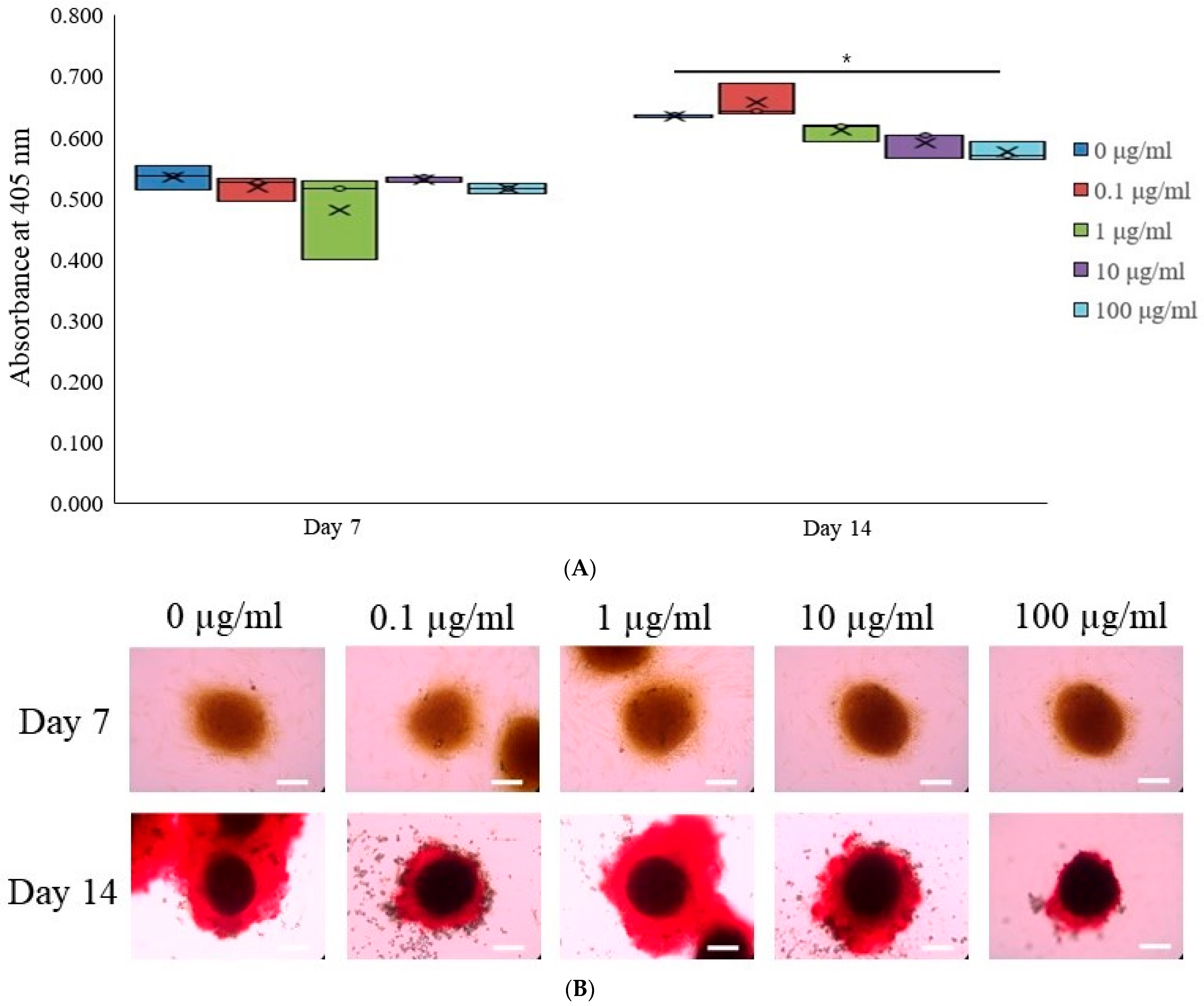
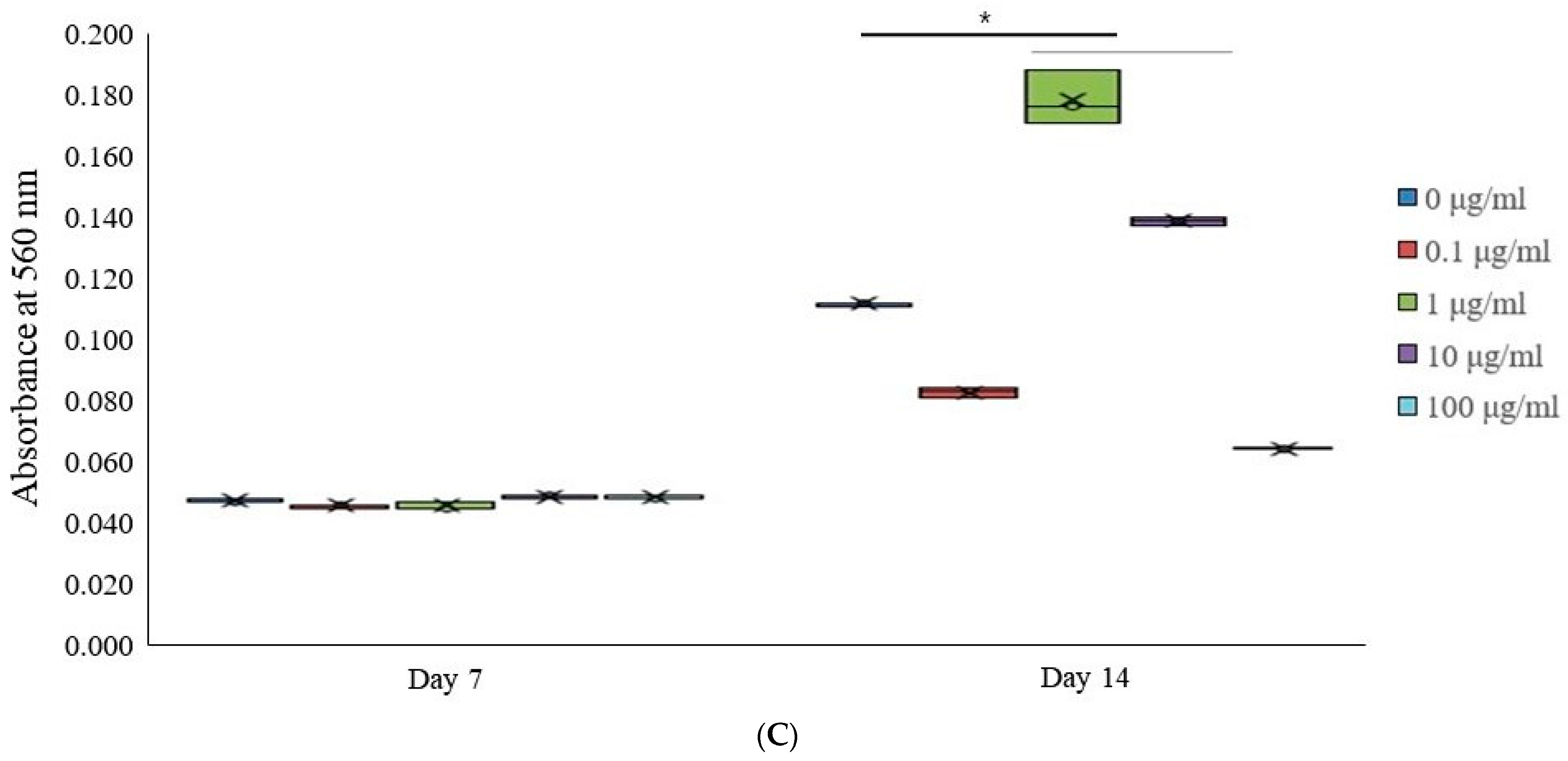
3.3. Alkaline Phosphatase Activity Levels and Calcium Deposition Extent
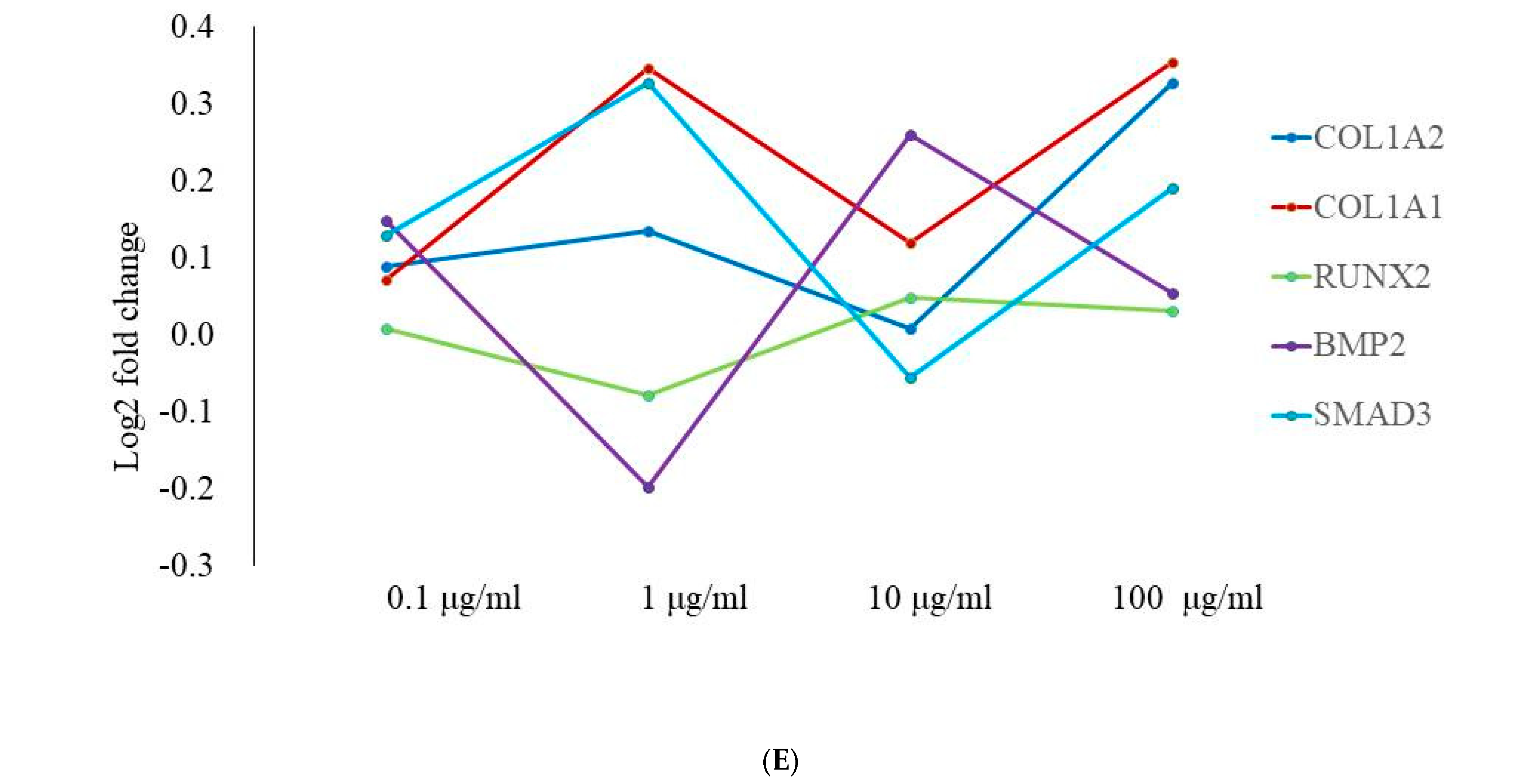
3.4. Total RNA Extraction and Real-Time Polymerase Chain Reaction Quantification
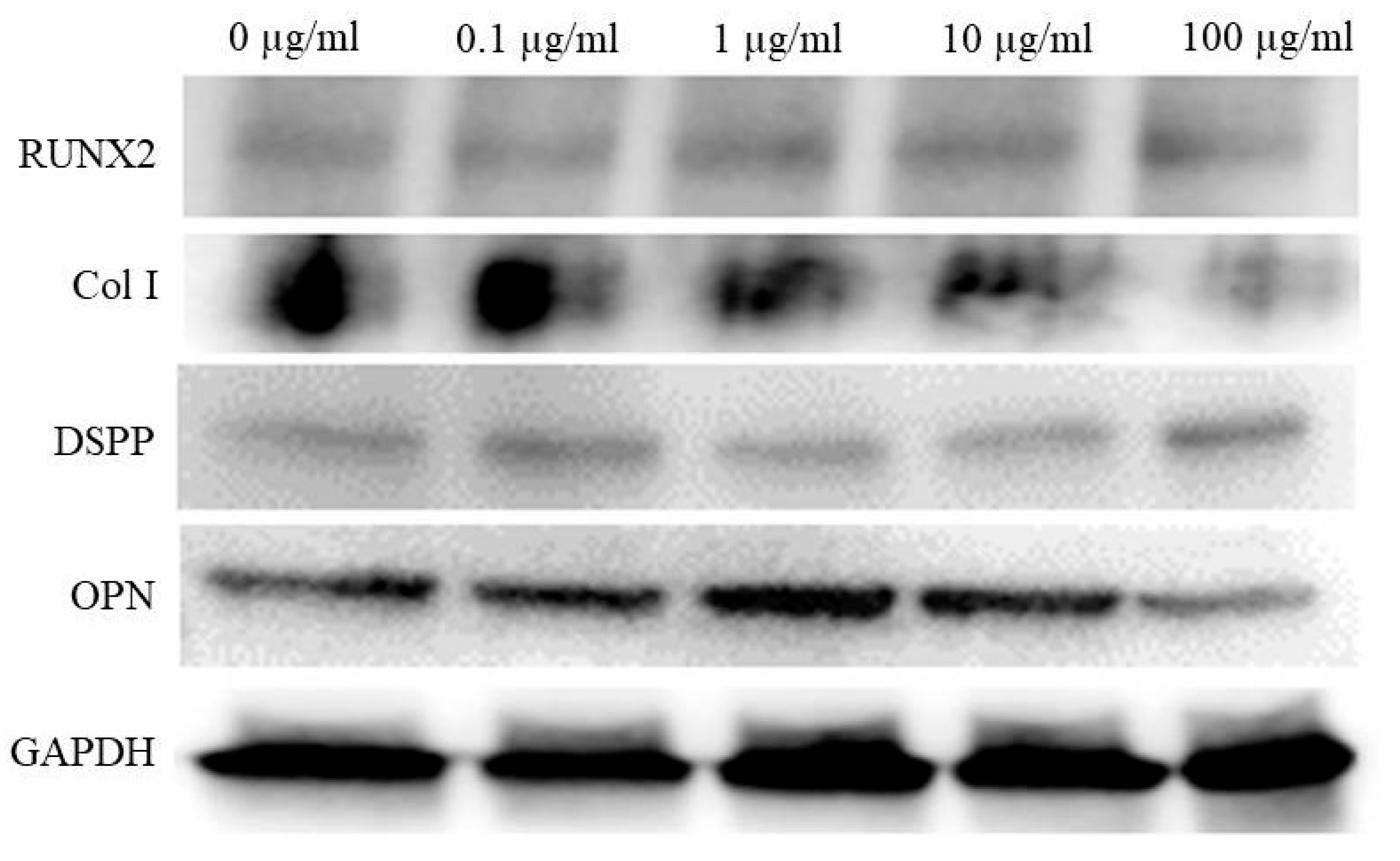
3.5. Western Blot Analysis
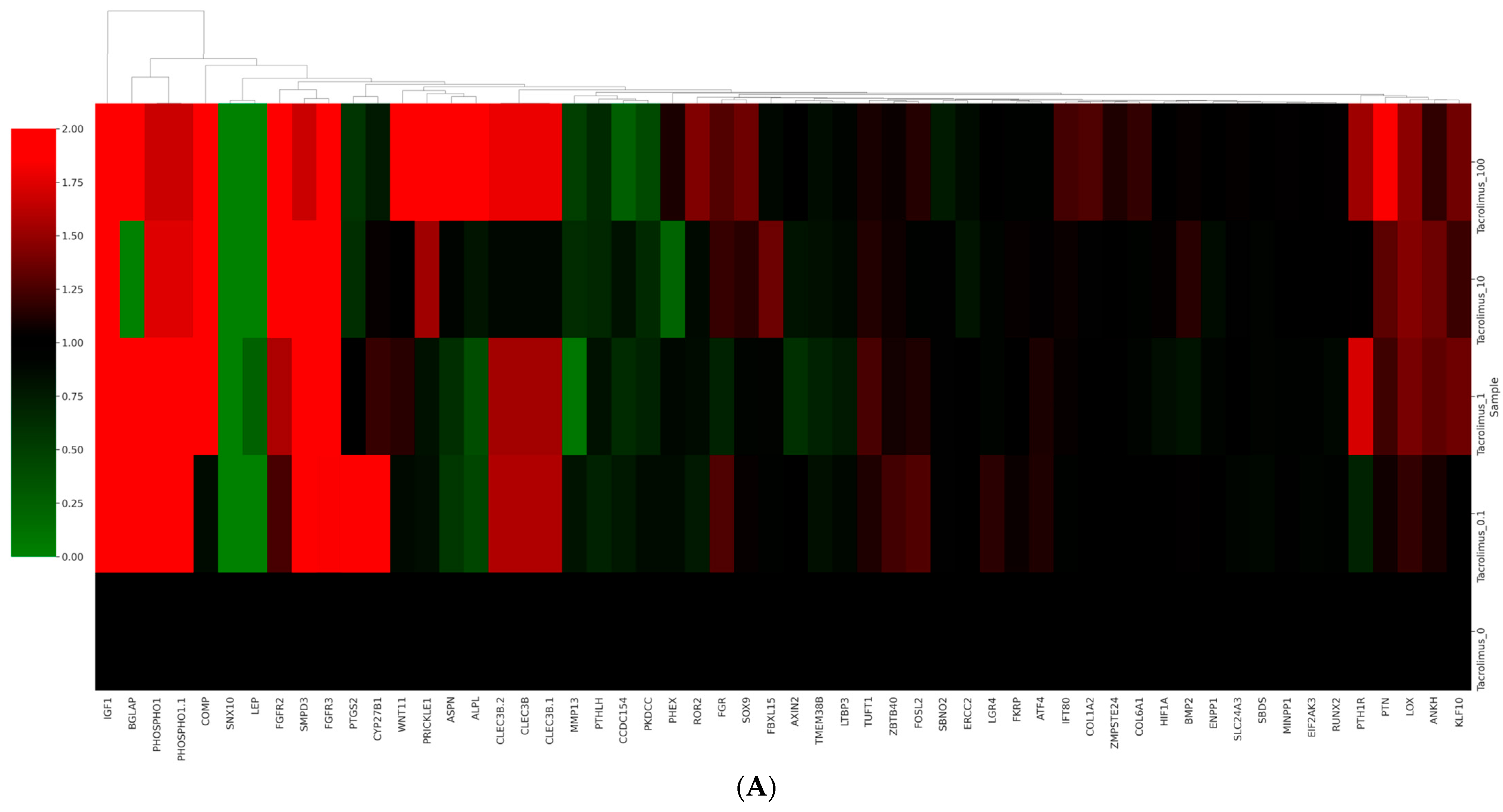
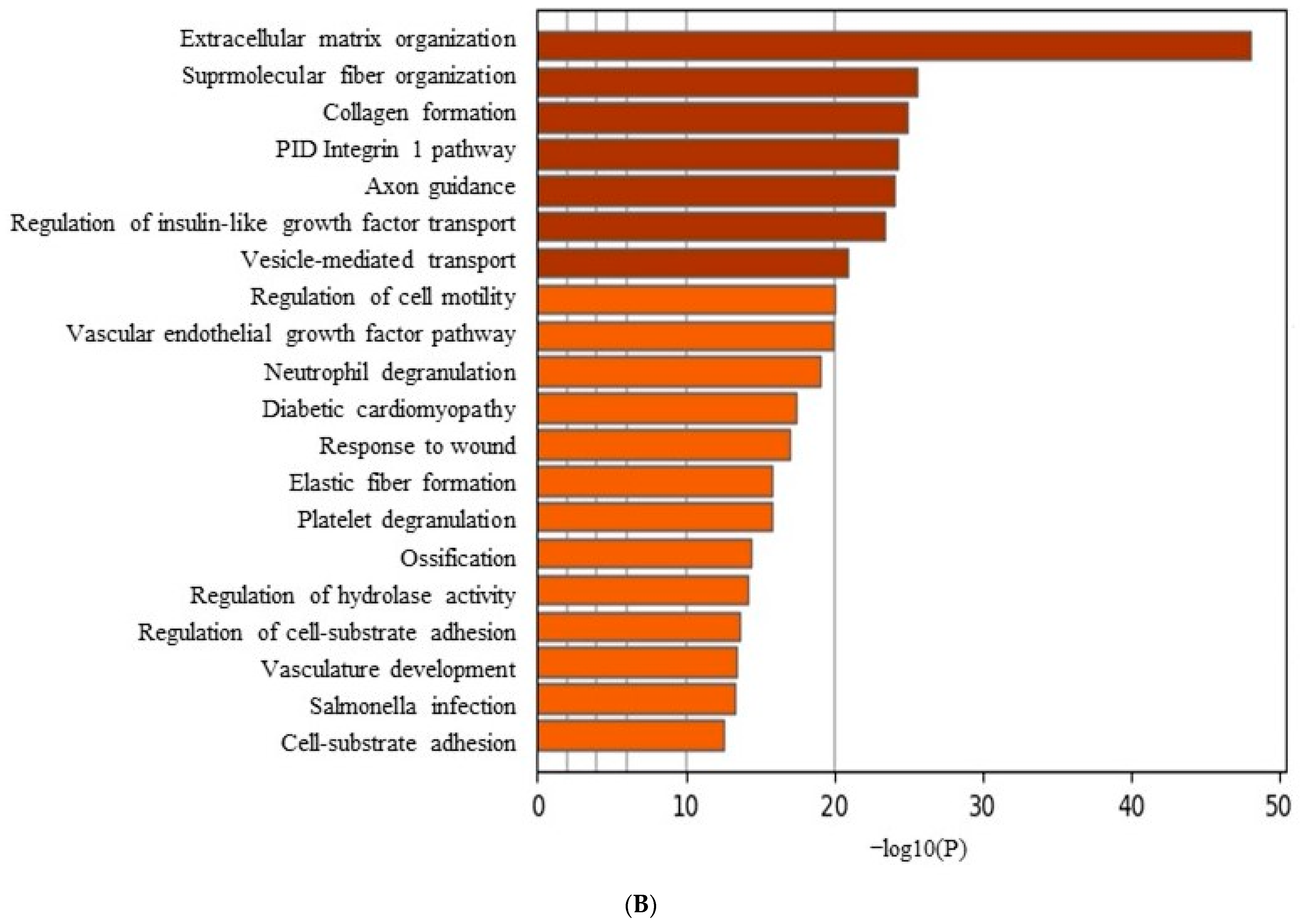
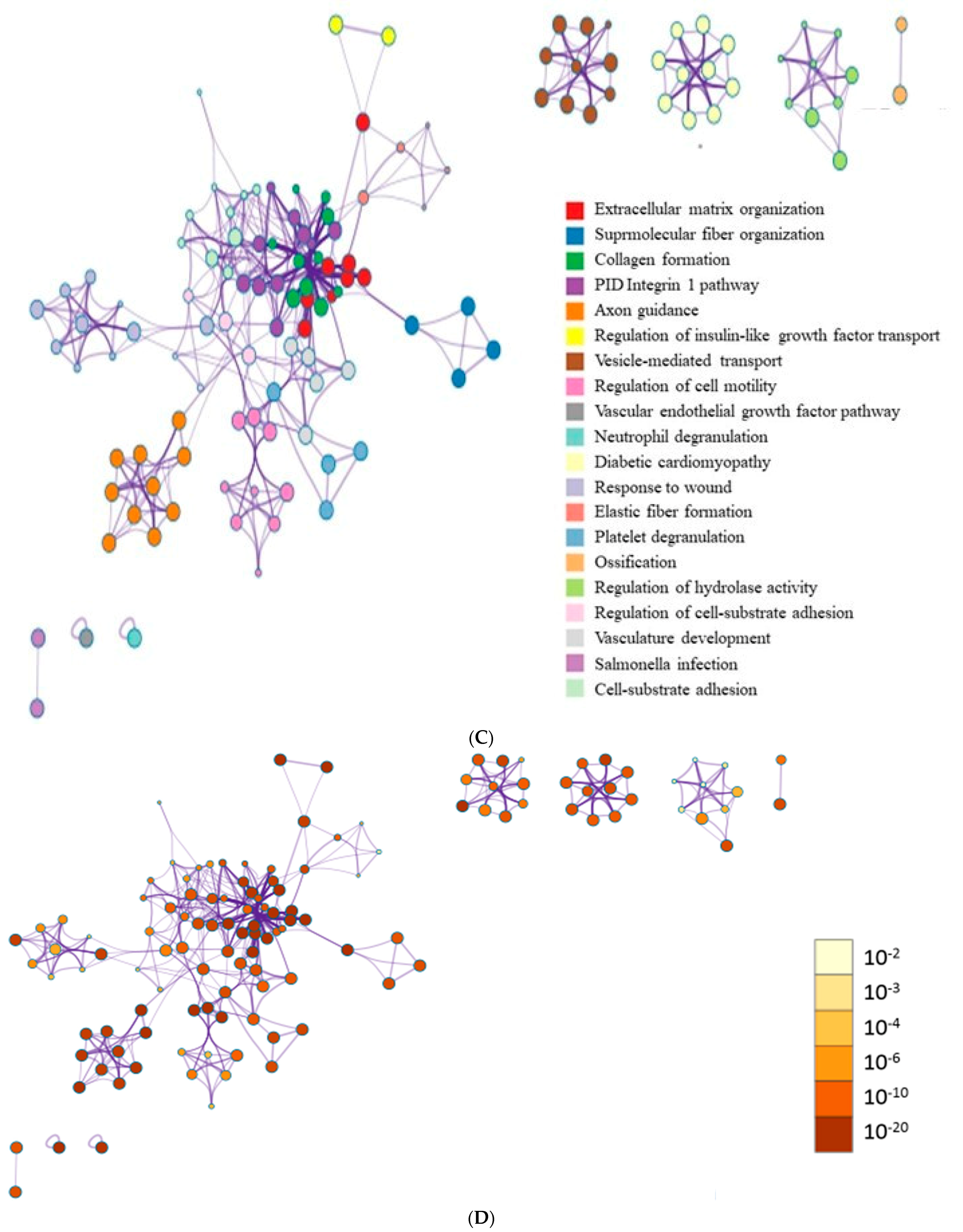
3.6. Data Analysis of RNA Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turło, J.; Gajzlerska, W.; Klimaszewska, M.; Król, M.; Dawidowski, M.; Gutkowska, B. Enhancement of tacrolimus productivity in Streptomyces tsukubaensis by the use of novel precursors for biosynthesis. Enzym. Microb. Technol. 2012, 51, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.C.; Gaston, R.S. Thirty Years of Tacrolimus in Clinical Practice. Transplantation 2021, 105, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liu, Y.; Yang, X.; He, J.; Zhang, F.; Zhong, Q.; Guo, X. Strontium ranelate promotes chondrogenesis through inhibition of the Wnt/β-catenin pathway. Stem Cell Res. Ther. 2021, 12, 296. [Google Scholar] [CrossRef] [PubMed]
- Chatterjea, A.; LaPointe, V.L.; Alblas, J.; Chatterjea, S.; van Blitterswijk, C.A.; de Boer, J. Suppression of the immune system as a critical step for bone formation from allogeneic osteoprogenitors implanted in rats. J. Cell. Mol. Med. 2014, 18, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Farmer, J.D., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Assmann, T.; Homey, B.; Ruzicka, T. Applications of tacrolimus for the treatment of skin disorders. Immunopharmacology 2000, 47, 203–213. [Google Scholar] [CrossRef]
- Ha, D.H.; Yong, C.S.; Kim, J.O.; Jeong, J.H.; Park, J.B. Effects of tacrolimus on morphology, proliferation and differentiation of mesenchymal stem cells derived from gingiva tissue. Mol. Med. Rep. 2016, 14, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.H.; Pathak, S.; Yong, C.S.; Kim, J.O.; Jeong, J.H.; Park, J.B. Potential differentiation ability of gingiva originated human mesenchymal stem cell in the presence of tacrolimus. Sci. Rep. 2016, 6, 34910. [Google Scholar] [CrossRef] [PubMed]
- Resende, J.P.; Chaves, M.; Aarestrup, F.M.; Aarestrup, B.V.; Olate, S.; Netto, H.D. Oral lichen planus treated with tacrolimus 0.1%. Int. J. Clin. Exp. Med. 2013, 6, 917–921. [Google Scholar]
- Riano Arguelles, A.; Martino Gorbea, R.; Iglesias Zamora, M.E.; Garatea Crelgo, J. Topic tacrolimus, alternative treatment for oral erosive lichen planus resistant to steroids: A case report. Med. Oral Patol. Oral Cir. Bucal 2006, 11, E462–E466. [Google Scholar]
- Nivethitha, K.; Ramesh, A.; Talwar, A.; Shenoy, N. Rare phenomena of tacrolimus-induced gingival hyperplasia. J. Oral Maxillofac. Pathol. JOMFP 2020, 24, 403. [Google Scholar] [PubMed]
- Li, L.; Neaves, W.B. Normal stem cells and cancer stem cells: The niche matters. Cancer Res. 2006, 66, 4553–4557. [Google Scholar] [CrossRef]
- Lee, D.H.; Yun, D.W.; Kim, Y.H.; Im, G.B.; Hyun, J.; Park, H.S.; Bhang, S.H.; Choi, S.H. Various Three-Dimensional Culture Methods and Cell Types for Exosome Production. Tissue Eng. Regen. Med. 2023, 20, 621–635. [Google Scholar] [CrossRef]
- Park, J.B.; Kim, I.; Lee, W.; Kim, H. Evaluation of the regenerative capacity of stem cells combined with bone graft material and collagen matrix using a rabbit calvarial defect model. J. Periodontal Implant Sci. 2023, 53, 467–477. [Google Scholar] [CrossRef]
- Sun, Q.; Nakata, H.; Yamamoto, M.; Kasugai, S.; Kuroda, S. Comparison of gingiva-derived and bone marrow mesenchymal stem cells for osteogenesis. J. Cell. Mol. Med. 2019, 23, 7592–7601. [Google Scholar] [CrossRef]
- Grawish, M.E. Gingival-derived mesenchymal stem cells: An endless resource for regenerative dentistry. World J. Stem Cells 2018, 10, 116–118. [Google Scholar] [CrossRef]
- Jin, S.H.; Lee, J.E.; Yun, J.H.; Kim, I.; Ko, Y.; Park, J.B. Isolation and characterization of human mesenchymal stem cells from gingival connective tissue. J. Periodontal Res. 2015, 50, 461–467. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, H.; Zhang, X.; Mao, T.; Lu, Y.; Gu, Y.; Ju, D.; Qi, L.; Wang, Q.; Dong, C. Construction of a decellularized spinal cord matrix/GelMA composite scaffold and its effects on neuronal differentiation of neural stem cells. J. Biomater. Sci. Polym. Ed. 2022, 33, 2124–2144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Y.; Zhang, Y.; Liu, Z.; Zhao, X. YTHDF1 promotes the viability and self-renewal of glioma stem cells by enhancing LINC00900 stability. Int. J. Oncol. 2024, 64, 53. [Google Scholar] [CrossRef]
- Raik, S.; Thakur, R.; Rattan, V.; Kumar, N.; Pal, A.; Bhattacharyya, S. Temporal Modulation of DNA Methylation and Gene Expression in Monolayer and 3D Spheroids of Dental Pulp Stem Cells during Osteogenic Differentiation: A Comparative Study. Tissue Eng. Regen. Med. 2022, 19, 1267–1282. [Google Scholar] [CrossRef]
- Yang, G.; Liu, K.; Ma, S.; Qi, P. PPARγ inhibition promotes osteogenic differentiation of bone marrow mesenchymal stem cells and fracture healing. J. Cell. Biochem. 2024. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Yu, Y.; Park, G.C.; Shin, S.C.; Kim, J.M.; Lee, B.J.; Kim, Y.H. Proliferation-Related Features of the Human Mesenchymal Stem Cells Derived from Palatine Tonsils, Adipose Tissues, and Bone Marrow. Tissue Eng. Regen. Med. 2023, 20, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Fu, S.; Zhang, H.; Li, X.; Zhang, Q.; Guo, C.; Qiu, K.; Feng, J.; Liu, X.; Liu, D. Exosomes Derived from Human Amniotic Mesenchymal Stem Cells Facilitate Diabetic Wound Healing by Angiogenesis and Enrich Multiple lncRNAs. Tissue Eng. Regen. Med. 2023, 20, 295–308. [Google Scholar] [CrossRef]
- Ertosun, M.G.; Özkan, Ö.; Özkan, Ö. Effects of Different Tacrolimus Doses on Adipose-Derived Stem Cells. Exp. Clin. Transplant. 2021, 19, 723–731. [Google Scholar] [CrossRef]
- Tsuji, W.; Schnider, J.T.; McLaughlin, M.M.; Schweizer, R.; Zhang, W.; Solari, M.G.; Rubin, J.P.; Marra, K.G.; Plock, J.A.; Gorantla, V.S. Effects of immunosuppressive drugs on viability and susceptibility of adipose- and bone marrow-derived mesenchymal stem cells. Front. Immunol. 2015, 6, 131. [Google Scholar] [CrossRef]
- Cayami, F.K.; Claeys, L.; de Ruiter, R.; Smilde, B.J.; Wisse, L.; Bogunovic, N.; Riesebos, E.; Eken, L.; Kooi, I.; Sistermans, E.A.; et al. Osteogenic transdifferentiation of primary human fibroblasts to osteoblast-like cells with human platelet lysate. Sci. Rep. 2022, 12, 14686. [Google Scholar] [CrossRef]
- Choi, S.W.; Seo, S.; Hong, H.K.; Yoon, S.J.; Kim, M.; Moon, S.; Lee, J.Y.; Lim, J.; Lee, J.B.; Woo, S.J. Therapeutic Extracellular Vesicles from Tonsil-Derived Mesenchymal Stem Cells for the Treatment of Retinal Degenerative Disease. Tissue Eng. Regen. Med. 2023, 20, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Hoogduijn, M.J.; Crop, M.J.; Korevaar, S.S.; Peeters, A.M.; Eijken, M.; Maat, L.P.; Balk, A.H.; Weimar, W.; Baan, C.C. Susceptibility of human mesenchymal stem cells to tacrolimus, mycophenolic acid, and rapamycin. Transplantation 2008, 86, 1283–1291. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, W.J.; Ryoo, H.M. Post-Translational Regulations of Transcriptional Activity of RUNX2. Mol. Cells 2020, 43, 160–167. [Google Scholar]
- Hia, E.M.; Jang, S.R.; Maharjan, B.; Park, J.; Park, C.H.; Kim, C.S. Construction of a PEGDA/chitosan hydrogel incorporating mineralized copper-doped mesoporous silica nanospheres for accelerated bone regeneration. Int. J. Biol. Macromol. 2024, 262, 130218. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Ghosh, J.; Dhara, S.K. Osteogenic differentiation potential of porcine bone marrow mesenchymal stem cell subpopulations selected in different basal media. Biol. Open 2020, 9, bio053280. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, S.; Gao, Q.; Kotecha, M.; Magin, R.L.; Karol, S.; Bedran-Russo, A.; George, A. Biomimetic extracellular matrix-incorporated scaffold induces osteogenic gene expression in human marrow stromal cells. Tissue Eng. Part A 2012, 18, 295–309. [Google Scholar] [CrossRef]
- Devos, H.; Zoidakis, J.; Roubelakis, M.G.; Latosinska, A.; Vlahou, A. Reviewing the Regulators of COL1A1. Int. J. Mol. Sci. 2023, 24, 10004. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, C.; Bai, H.; Zhang, J.; Wang, Z.; Li, Z.; Zhao, X.; Wang, J.; Liu, H. Functionalization of biomimetic mineralized collagen for bone tissue engineering. Mater. Today Bio 2023, 20, 100660. [Google Scholar] [CrossRef]
- Arai, Y.; Lee, S.H. MMP13-Overexpressing Mesenchymal Stem Cells Enhance Bone Tissue Formation in the Presence of Collagen Hydrogel. Tissue Eng. Regen. Med. 2023, 20, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E. Comparison of conventional and advanced in vitro models in the toxicity testing of nanoparticles. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1091–1107. [Google Scholar] [CrossRef]
- Gastaca, M.; Ruiz, P.; Bustamante, J.; Martinez-Indart, L.; Ventoso, A.; Fernandez, J.R.; Palomares, I.; Prieto, M.; Testillano, M.; Salvador, P.; et al. Early tacrolimus exposure does not impact long-term outcomes after liver transplantation. World J. Hepatol. 2021, 13, 362–374. [Google Scholar] [CrossRef]
- Wilson, W.E.; Suh, H.H. Fine particles and coarse particles: Concentration relationships relevant to epidemiologic studies. J. Air Waste Manag. Assoc. 1997, 47, 1238–1249. [Google Scholar] [CrossRef]
- Sontag, C.J.; Nguyen, H.X.; Kamei, N.; Uchida, N.; Anderson, A.J.; Cummings, B.J. Immunosuppressants affect human neural stem cells in vitro but not in an in vivo model of spinal cord injury. Stem Cells Transl. Med. 2013, 2, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Penny, T.; Pham, Y.; Sutherland, A.E.; Jenkin, G.; Fahey, M.C.; Paton, M.C.B.; Finch-Edmondson, M.; Miller, S.L.; McDonald, C.A. Neuroprotective Action of Tacrolimus before and after Onset of Neonatal Hypoxic-Ischaemic Brain Injury in Rats. Cells 2023, 12, 2659. [Google Scholar] [CrossRef] [PubMed]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, W.-J.; Han, S.-H.; Lee, H.-J.; Kim, J.-H.; Song, H.-J.; Park, J.-B. The Influence of Tacrolimus on Cellular Morphology, Cellular Viability, Osteogenic Differentiation, and mRNA Expression within Stem Cell Spheroids. Medicina 2024, 60, 702. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina60050702
Park W-J, Han S-H, Lee H-J, Kim J-H, Song H-J, Park J-B. The Influence of Tacrolimus on Cellular Morphology, Cellular Viability, Osteogenic Differentiation, and mRNA Expression within Stem Cell Spheroids. Medicina. 2024; 60(5):702. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina60050702
Chicago/Turabian StylePark, Won-Jong, Sung-Hoon Han, Hyun-Jin Lee, Ju-Hwan Kim, Hye-Jung Song, and Jun-Beom Park. 2024. "The Influence of Tacrolimus on Cellular Morphology, Cellular Viability, Osteogenic Differentiation, and mRNA Expression within Stem Cell Spheroids" Medicina 60, no. 5: 702. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina60050702