

The Effect of Gut Microbiota-Targeted Interventions on Neuroinflammation and Motor Function in Parkinson’s Disease Animal Models—A Systematic Review
 , and
, and
Abstract
:1. Introduction
1.1. Parkinson’s Disease—Clinical and Pathophysiological Characterization
1.2. The Gut–Brain Axis and Its Role in PD Pathogenesis
1.3. The Role of the Gut Microbiota in PD Pathogenesis
1.4. CNS and GIT Inflammatory Biomarkers in PD
1.5. Non-Dietary Gut Microbiota-Targeted Interventions
1.6. Clinical Translation of Rodent Studies on Microbiota-Targeted Therapies in PD
2. Materials and Methods
2.1. Search Strategy and Protocol Registration
- PubMed: (“parkinsonian disorders”[MeSH Terms] OR (“parkinsonian”[All Fields] AND “disorders”[All Fields]) OR “parkinsonian disorders”[All Fields]) AND ((“fecal microbiota transplantation”[MeSH Terms] OR (“fecal”[All Fields] AND “microbiota”[All Fields] AND “transplantation”[All Fields]) OR “fecal microbiota transplantation”[All Fields]) OR (“probiotics"[MeSH Terms] OR “probiotics”[All Fields]) OR (“prebiotics”[MeSH Terms] OR “prebiotics”[All Fields]) OR (“anti-bacterial agents”[All Fields] OR “anti-bacterial agents”[MeSH Terms] OR (“anti-bacterial”[All Fields] AND “agents"[All Fields]) OR “anti-bacterial agents”[All Fields] OR “antibiotics”[All Fields]));
- SCOPUS: TITLE-ABS-KEY (neuroinflammation AND (Parkinson AND disease) AND ((fecal AND microbiota AND transplantation) OR probiotics OR prebiotics OR (anti-bacterial AND agents) OR antibiotics));
- Embase: (parkinsonism)/br AND ((‘fecal microbiota transplantation’)/br) OR ((‘probiotic agent’):ti) OR ((‘prebiotic agent’):ti) OR ((‘antiinfective agent’):ti) OR ((‘antibiotic agent’):ti);
- Web of Science: Parkinson’s AND (fecal microbiota transplantation OR (fecal AND microbiota AND transplantation) OR probiotics OR prebiotics OR anti-bacterial agents OR (anti-bacterial AND agents) OR antibiotics).
2.2. PRISMA Guidelines
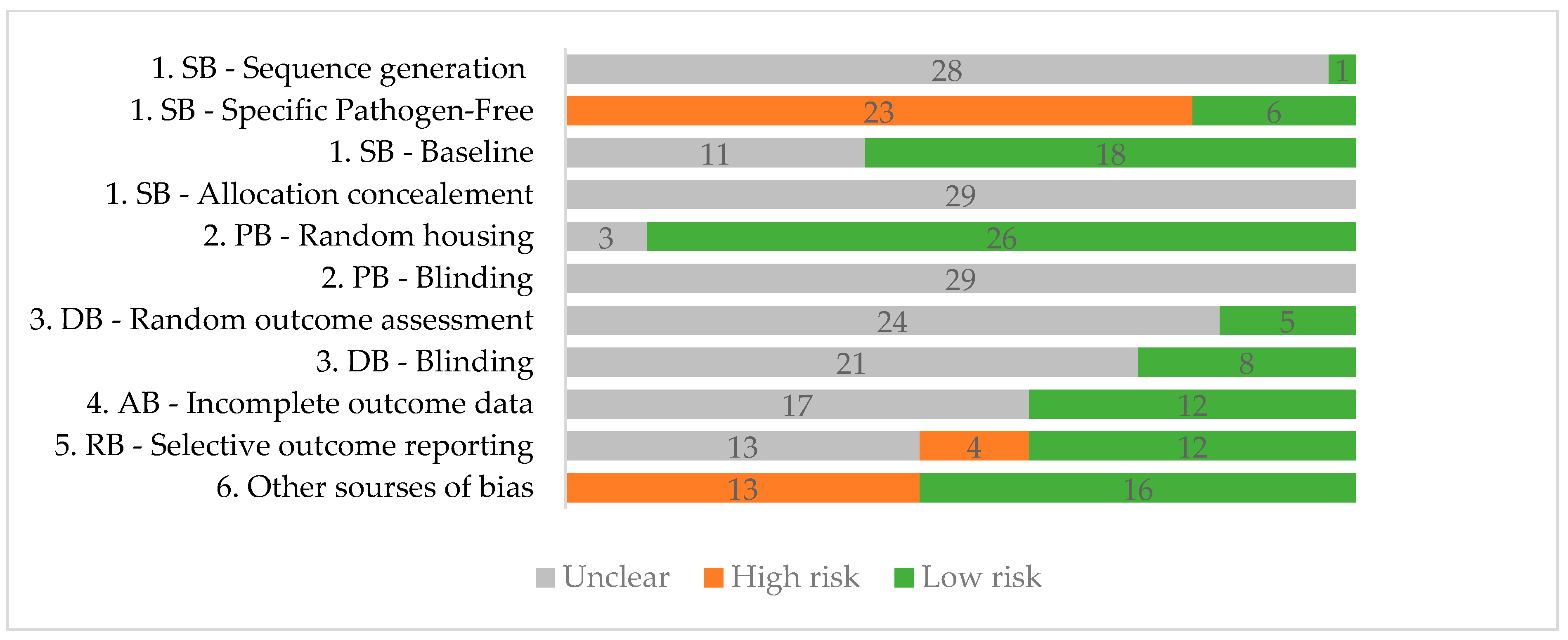
2.3. SYRCLE Risk of Bias
2.4. Synthesis without Meta-Analysis (SWiM)
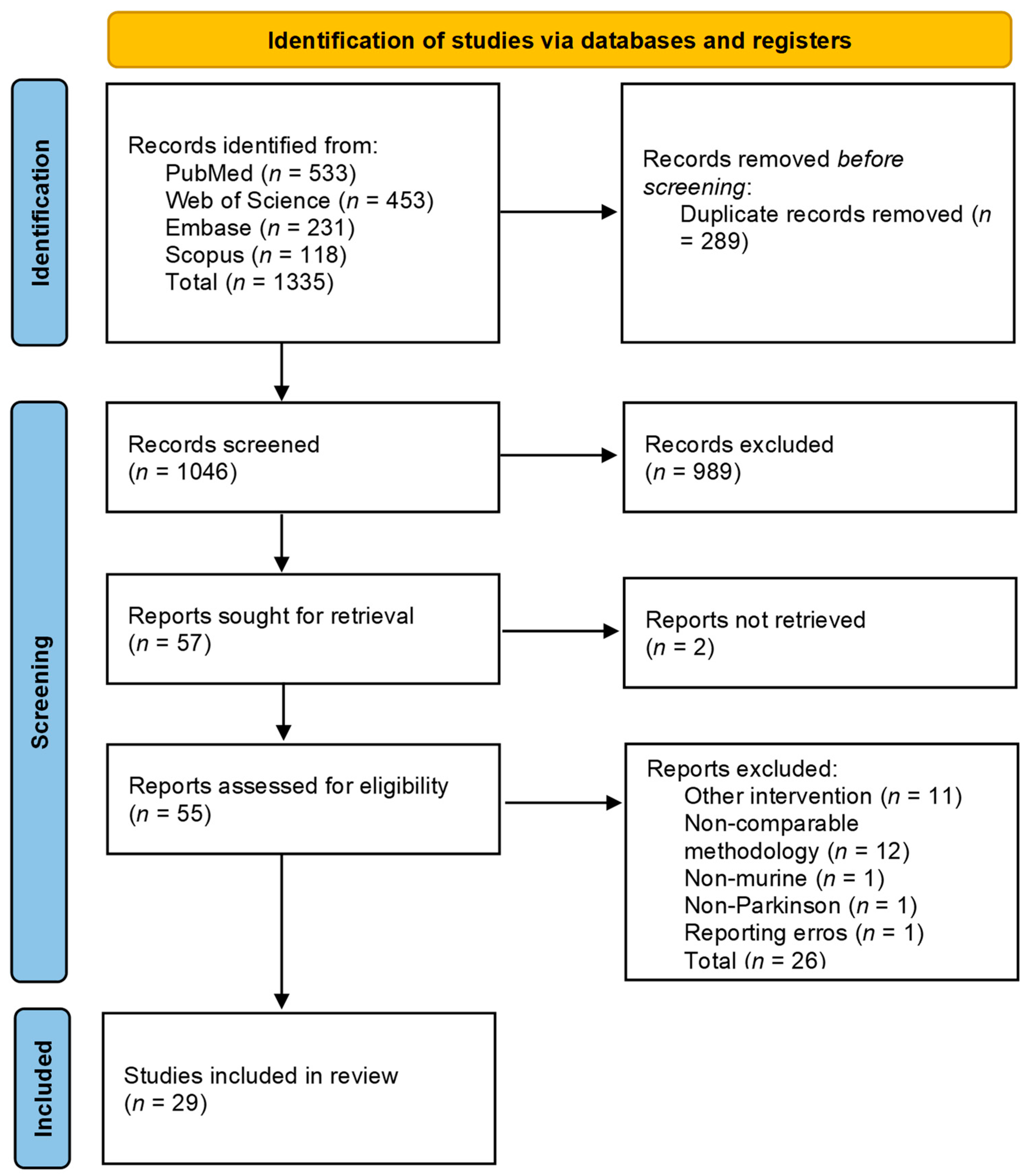
3. Results
3.1. PRISMA Flowchart
3.2. Quality Assessment
3.3. Studies’ Designs
3.4. Primary Outcomes
3.4.1. Dopaminergic Cell Loss
3.4.2. Microglial Activation
3.4.3. Astrocytic Activation
3.4.4. Cytokines
3.4.5. Behavioral Tests
3.4.6. Microbiome Relative Abundance
3.5. Secondary Outcomes
4. Discussion
4.1. Metodological Considerations
4.2. Main Findings
4.3. Future Research
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2019 Dementia Collaborators. The Burden of Dementia Due to Down Syndrome, Parkinson’s Disease, Stroke, and Traumatic Brain Injury: A Systematic Analysis for the Global Burden of Disease Study 2019. Neuroepidemiology 2021, 55, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, E.R.; Sherer, T.; Okun, M.S.; Bloem, B.R. The Emerging Evidence of the Parkinson Pandemic. J. Park. Dis. 2018, 8, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulos, I.C.; Malli, F.; Sinani, O.; Gourgoulianis, K.I.; Xiromerisiou, G. Worldwide Trends in Mortality Related to Parkinson’s Disease in the Period of 1994-2019: Analysis of Vital Registration Data from the WHO Mortality Database. Front. Neurol. 2022, 13, 956440. [Google Scholar] [CrossRef] [PubMed]
- Pinter, B.; Diem-Zangerl, A.; Wenning, G.K.; Scherfler, C.; Oberaigner, W.; Seppi, K.; Poewe, W. Mortality in Parkinson’s Disease: A 38-year Follow-up Study. Mov. Disord. 2015, 30, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.K.; Tanner, C.M.; Brundin, P. Parkinson Disease Epidemiology, Pathology, Genetics, and Pathophysiology. Clin. Geriatr. Med. 2020, 36, 1–12. [Google Scholar] [CrossRef]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s Disease. Lancet Lond. Engl. 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J. Determinants of Dopaminergic Neuron Loss in Parkinson’s Disease. FEBS J. 2018, 285, 3657–3668. [Google Scholar] [CrossRef] [PubMed]
- Morris, H.R.; Spillantini, M.G.; Sue, C.M.; Williams-Gray, C.H. The Pathogenesis of Parkinson’s Disease. Lancet Lond. Engl. 2024, 403, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Liddle, R.A. Parkinson’s Disease from the Gut. Brain Res. 2018, 1693, 201–206. [Google Scholar] [CrossRef]
- Burré, J.; Sharma, M.; Südhof, T.C. Cell Biology and Pathophysiology of α-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8, a024091. [Google Scholar] [CrossRef]
- Costa, H.N.; Esteves, A.R.; Empadinhas, N.; Cardoso, S.M. Parkinson’s Disease: A Multisystem Disorder. Neurosci. Bull. 2023, 39, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Hey, G.; Nair, N.; Klann, E.; Gurrala, A.; Safarpour, D.; Mai, V.; Ramirez-Zamora, A.; Vedam-Mai, V. Therapies for Parkinson’s Disease and the Gut Microbiome: Evidence for Bidirectional Connection. Front. Aging Neurosci. 2023, 15, 1151850. [Google Scholar] [CrossRef] [PubMed]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Yemula, N.; Dietrich, C.; Dostal, V.; Hornberger, M. Parkinson’s Disease and the Gut: Symptoms, Nutrition, and Microbiota. J. Park. Dis. 2021, 11, 1491–1505. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.; Ahmad, F.; Banu, A.; Mohammad, F. Gut Microbiome and Parkinson’s Disease: Perspective on Pathogenesis and Treatment. J. Adv. Res. 2023, 50, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Seguella, L.; Sarnelli, G.; Esposito, G. Leaky Gut, Dysbiosis, and Enteric Glia Activation: The Trilogy behind the Intestinal Origin of Parkinson’s Disease. Neural Regen. Res. 2020, 15, 1037–1038. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-B.; Zhao, D.-Y.; Qi, Q.-Q.; Long, X.; Li, X.; Chen, F.-X.; Zuo, X.-L. BDNF Modulates Intestinal Barrier Integrity through Regulating the Expression of Tight Junction Proteins. Neurogastroenterol. Motil. 2017, 29, 12967. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cai, Y.-Y.; Yan, Z.-X. Brain-Derived Neurotrophic Factor Preserves Intestinal Mucosal Barrier Function and Alters Gut Microbiota in Mice. Kaohsiung J. Med. Sci. 2018, 34, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, N.; Zhang, Z.; Li, H.; Ma, J.; Yuan, Y.; Shi, M.; Liu, J.; Zhao, Z.; Zhang, Z.; et al. Cholecystokinin and Glucagon-like Peptide-1 Analogues Regulate Intestinal Tight Junction, Inflammation, Dopaminergic Neurons and α-Synuclein Accumulation in the Colon of Two Parkinson’s Disease Mouse Models. Eur. J. Pharmacol. 2022, 926, 175029. [Google Scholar] [CrossRef]
- Wallen, Z.D.; Demirkan, A.; Twa, G.; Cohen, G.; Dean, M.N.; Standaert, D.G.; Sampson, T.R.; Payami, H. Metagenomics of Parkinson’s Disease Implicates the Gut Microbiome in Multiple Disease Mechanisms. Nat. Commun. 2022, 13, 6958. [Google Scholar] [CrossRef]
- Dos Santos, J.C.C.; Oliveira, L.F.; Noleto, F.M.; Gusmão, C.T.P.; de Castro Brito, G.A.; de Barros Viana, G.S. Gut-Microbiome-Brain Axis: The Crosstalk between the Vagus Nerve, Alpha-Synuclein and the Brain in Parkinson’s Disease. Neural Regen. Res. 2023, 18, 2611–2614. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Meng, L.-B.; Chen, L.-J.; Shi, X.; Tu, L.; Zhou, Q.; Yu, J.-L.; Liao, X.; Zeng, Y.; Yuan, Q.-Y. The Role of the Microbiota-Gut-Brain Axis and Intestinal Microbiome Dysregulation in Parkinson’s Disease. Front. Neurol. 2023, 14, 1185375. [Google Scholar] [CrossRef] [PubMed]
- Klann, E.M.; Dissanayake, U.; Gurrala, A.; Farrer, M.; Shukla, A.W.; Ramirez-Zamora, A.; Mai, V.; Vedam-Mai, V. The Gut-Brain Axis and Its Relation to Parkinson’s Disease: A Review. Front. Aging Neurosci. 2021, 13, 782082. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lu, J.; Wei, K.; Wei, J.; Tian, P.; Yue, M.; Wang, Y.; Hong, D.; Li, F.; Wang, B.; et al. Neuroprotective Effect of Ceftriaxone on MPTP-Induced Parkinson’s Disease Mouse Model by Regulating Inflammation and Intestinal Microbiota. Oxid. Med. Cell. Longev. 2021, 2021, 9424582. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Zhu, W.; Hosoda, W.; Sen, J.M.; Mattson, M.P. Chronic Mild Gut Inflammation Accelerates Brain Neuropathology and Motor Dysfunction in α-Synuclein Mutant Mice. Neuromol. Med. 2019, 21, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Hor, J.W.; Lim, S.-Y.; Khor, E.S.; Chong, K.K.; Song, S.L.; Ibrahim, N.M.; Teh, C.S.J.; Chong, C.W.; Hilmi, I.N.; Tan, A.H. Fecal Calprotectin in Parkinson’s Disease and Multiple System Atrophy. J. Mov. Disord. 2022, 15, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Spiegel, J.; Dillmann, U.; Grundmann, D.; Bürmann, J.; Faßbender, K.; Schäfer, K.-H.; Unger, M.M. Fecal Markers of Intestinal Inflammation and Intestinal Permeability Are Elevated in Parkinson’s Disease. Parkinsonism Relat. Disord. 2018, 50, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; El-Hadidi, M.; Admard, J.; Wassouf, Z.; Schulze-Hentrich, J.M.; Kohlhofer, U.; Quintanilla-Martinez, L.; Huson, D.; Riess, O.; Casadei, N. Enriched Environmental Conditions Modify the Gut Microbiome Composition and Fecal Markers of Inflammation in Parkinson’s Disease. Front. Neurosci. 2019, 13, 1032. [Google Scholar] [CrossRef]
- Lehmann, F.S.; Burri, E.; Beglinger, C. The Role and Utility of Faecal Markers in Inflammatory Bowel Disease. Ther. Adv. Gastroenterol. 2015, 8, 23–36. [Google Scholar] [CrossRef]
- Houser, M.C.; Chang, J.; Factor, S.A.; Molho, E.S.; Zabetian, C.P.; Hill-Burns, E.M.; Payami, H.; Hertzberg, V.S.; Tansey, M.G. Stool Immune Profiles Evince Gastrointestinal Inflammation in Parkinson’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2018, 33, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Dumitrescu, L.; Marta, D.; Dănău, A.; Lefter, A.; Tulbă, D.; Cozma, L.; Manole, E.; Gherghiceanu, M.; Ceafalan, L.C.; Popescu, B.O. Serum and Fecal Markers of Intestinal Inflammation and Intestinal Barrier Permeability Are Elevated in Parkinson’s Disease. Front. Neurosci. 2021, 15, 689723. [Google Scholar] [CrossRef]
- Bicknell, B.; Liebert, A.; Borody, T.; Herkes, G.; McLachlan, C.; Kiat, H. Neurodegenerative and Neurodevelopmental Diseases and the Gut-Brain Axis: The Potential of Therapeutic Targeting of the Microbiome. Int. J. Mol. Sci. 2023, 24, 9577. [Google Scholar] [CrossRef]
- Bartl, M.; Dakna, M.; Schade, S.; Otte, B.; Wicke, T.; Lang, E.; Starke, M.; Ebentheuer, J.; Weber, S.; Toischer, K.; et al. Blood Markers of Inflammation, Neurodegeneration, and Cardiovascular Risk in Early Parkinson’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2023, 38, 68–81. [Google Scholar] [CrossRef]
- Qu, Y.; Li, J.; Qin, Q.; Wang, D.; Zhao, J.; An, K.; Mao, Z.; Min, Z.; Xiong, Y.; Li, J.; et al. A Systematic Review and Meta-Analysis of Inflammatory Biomarkers in Parkinson’s Disease. NPJ Park. Dis. 2023, 9, 18. [Google Scholar] [CrossRef]
- Xiromerisiou, G.; Marogianni, C.; Lampropoulos, I.C.; Dardiotis, E.; Speletas, M.; Ntavaroukas, P.; Androutsopoulou, A.; Kalala, F.; Grigoriadis, N.; Papoutsopoulou, S. Peripheral Inflammatory Markers TNF-α and CCL2 Revisited: Association with Parkinson’s Disease Severity. Int. J. Mol. Sci. 2022, 24, 264. [Google Scholar] [CrossRef]
- Kwon, E.H.; Tennagels, S.; Gold, R.; Gerwert, K.; Beyer, L.; Tönges, L. Update on CSF Biomarkers in Parkinson’s Disease. Biomolecules 2022, 12, 329. [Google Scholar] [CrossRef]
- Cacabelos, R. Parkinson’s Disease: From Pathogenesis to Pharmacogenomics. Int. J. Mol. Sci. 2017, 18, 551. [Google Scholar] [CrossRef] [PubMed]
- Elkouzi, A.; Vedam-Mai, V.; Eisinger, R.S.; Okun, M.S. Emerging Therapies in Parkinson Disease—Repurposed Drugs and New Approaches. Nat. Rev. Neurol. 2019, 15, 204–223. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, X.; Ye, Y.; Yan, X.; Cheng, Y.; Zhao, L.; Chen, F.; Ling, Z. Gut Microbiota: A Novel Therapeutic Target for Parkinson’s Disease. Front. Immunol. 2022, 13, 937555. [Google Scholar] [CrossRef] [PubMed]
- Alexoudi, A.; Kesidou, L.; Gatzonis, S.; Charalampopoulos, C.; Tsoga, A. Effectiveness of the Combination of Probiotic Supplementation on Motor Symptoms and Constipation in Parkinson’s Disease. Cureus 2023, 15, e49320. [Google Scholar] [CrossRef] [PubMed]
- Koutzoumis, D.N.; Vergara, M.; Pino, J.; Buddendorff, J.; Khoshbouei, H.; Mandel, R.J.; Torres, G.E. Alterations of the Gut Microbiota with Antibiotics Protects Dopamine Neuron Loss and Improve Motor Deficits in a Pharmacological Rodent Model of Parkinson’s Disease. Exp. Neurol. 2020, 325, 113159. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.-T.; Chan, L.; Chen, K.-Y.; Lee, H.-H.; Huang, L.-K.; Yang, Y.-C.S.H.; Liu, Y.-R.; Hu, C.-J. Rifaximin Modifies Gut Microbiota and Attenuates Inflammation in Parkinson’s Disease: Preclinical and Clinical Studies. Cells 2022, 11, 3468. [Google Scholar] [CrossRef] [PubMed]
- Matheson, J.-A.T.; Holsinger, R.M.D. The Role of Fecal Microbiota Transplantation in the Treatment of Neurodegenerative Diseases: A Review. Int. J. Mol. Sci. 2023, 24, 1001. [Google Scholar] [CrossRef] [PubMed]
- Vendrik, K.E.W.; Ooijevaar, R.E.; de Jong, P.R.C.; Laman, J.D.; van Oosten, B.W.; van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal Microbiota Transplantation in Neurological Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Segal, A.; Zlotnik, Y.; Moyal-Atias, K.; Abuhasira, R.; Ifergane, G. Fecal Microbiota Transplant as a Potential Treatment for Parkinson’s Disease—A Case Series. Clin. Neurol. Neurosurg. 2021, 207, 106791. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.-J.; Yang, X.-Z.; Tong, Q.; Shen, P.; Ma, S.-J.; Wu, S.-N.; Zheng, J.-L.; Wang, H.-G. Fecal Microbiota Transplantation Therapy for Parkinson’s Disease: A Preliminary Study. Medicine 2020, 99, e22035. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ning, J.; Bao, X.-Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal Microbiota Transplantation Protects Rotenone-Induced Parkinson’s Disease Mice via Suppressing Inflammation Mediated by the Lipopolysaccharide-TLR4 Signaling Pathway through the Microbiota-Gut-Brain Axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; Philippart de Foy, J.-M.; Dequenne, I.; de Timary, P.; Cani, P.D. How Probiotics Affect the Microbiota. Front. Cell. Infect. Microbiol. 2019, 9, 454. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, Prebiotics and Synbiotics: Safe Options for next-Generation Therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar] [CrossRef]
- Li, H.-Y.; Zhou, D.-D.; Gan, R.-Y.; Huang, S.-Y.; Zhao, C.-N.; Shang, A.; Xu, X.-Y.; Li, H.-B. Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review. Nutrients 2021, 13, 3211. [Google Scholar] [CrossRef]
- Tan, A.H.; Lim, S.-Y.; Chong, K.K.; A Manap, M.A.A.; Hor, J.W.; Lim, J.L.; Low, S.C.; Chong, C.W.; Mahadeva, S.; Lang, A.E. Probiotics for Constipation in Parkinson Disease: A Randomized Placebo-Controlled Study. Neurology 2021, 96, e772–e782. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Li, Y.; Xu, X.; Li, R.; Zhang, M.; Cui, Y.; Zhang, L.; Wei, Z.; Wang, S.; Tuo, H. Probiotics for Constipation and Gut Microbiota in Parkinson’s Disease. Parkinsonism Relat. Disord. 2022, 103, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Ali, R.A.R.; Manaf, M.R.A.; Ahmad, N.; Tajurruddin, F.W.; Qin, W.Z.; Desa, S.H.M.; Ibrahim, N.M. Multi-Strain Probiotics (Hexbio) Containing MCP BCMC Strains Improved Constipation and Gut Motility in Parkinson’s Disease: A Randomised Controlled Trial. PLoS ONE 2020, 15, e0244680. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; He, X.; Xu, S.; Zhang, Y.; Mo, C.; Lai, Y.; Song, Y.; Yan, Z.; Ai, P.; Qian, Y.; et al. Effect of Lacticaseibacillus Paracasei Strain Shirota Supplementation on Clinical Responses and Gut Microbiome in Parkinson’s Disease. Food Funct. 2023, 14, 6828–6839. [Google Scholar] [CrossRef] [PubMed]
- Tamtaji, O.R.; Taghizadeh, M.; Daneshvar Kakhaki, R.; Kouchaki, E.; Bahmani, F.; Borzabadi, S.; Oryan, S.; Mafi, A.; Asemi, Z. Clinical and Metabolic Response to Probiotic Administration in People with Parkinson’s Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Nutr. Edinb. Scotl. 2019, 38, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Qi, Y.; Chen, J.; Han, S.; Su, W.; Ma, X.; Yu, Y.; Wang, Y. Neuroprotective Effects of Bifidobacterium Animalis Subsp. Lactis NJ241 in a Mouse Model of Parkinson’s Disease: Implications for Gut Microbiota and PGC-1α. Mol. Neurobiol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Alli, S.R.; Gorbovskaya, I.; Liu, J.C.W.; Kolla, N.J.; Brown, L.; Müller, D.J. The Gut Microbiome in Depression and Potential Benefit of Prebiotics, Probiotics and Synbiotics: A Systematic Review of Clinical Trials and Observational Studies. Int. J. Mol. Sci. 2022, 23, 4494. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Boktor, J.C.; Sharon, G.; Verhagen Metman, L.A.; Hall, D.A.; Engen, P.A.; Zreloff, Z.; Hakim, D.J.; Bostick, J.W.; Ousey, J.; Lange, D.; et al. Integrated Multi-Cohort Analysis of the Parkinson’s Disease Gut Metagenome. Mov. Disord. Off. J. Mov. Disord. Soc. 2023, 38, 399–409. [Google Scholar] [CrossRef]
- Kleine Bardenhorst, S.; Cereda, E.; Severgnini, M.; Barichella, M.; Pezzoli, G.; Keshavarzian, A.; Desideri, A.; Pietrucci, D.; Aho, V.T.E.; Scheperjans, F.; et al. Gut Microbiota Dysbiosis in Parkinson Disease: A Systematic Review and Pooled Analysis. Eur. J. Neurol. 2023, 30, 3581–3594. [Google Scholar] [CrossRef]
- Hirayama, M.; Ohno, K. Parkinson’s Disease and Gut Microbiota. Ann. Nutr. Metab. 2021, 77 (Suppl. 2), 28–35. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Chen, D.; Zhu, X.; Cheng, C. Efficacy and Safety of Probiotics in Parkinson’s Constipation: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2022, 13, 1007654. [Google Scholar] [CrossRef] [PubMed]
- Van Laar, T.; Boertien, J.M.; Herranz, A.H. Faecal Transplantation, Pro- and Prebiotics in Parkinson’s Disease; Hope or Hype? J. Park. Dis. 2019, 9, S371–S379. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Zhu, F. Probiotics for Constipation in Parkinson’s: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Front. Cell. Infect. Microbiol. 2022, 12, 1038928. [Google Scholar] [CrossRef] [PubMed]
- Ghalandari, N.; Assarzadegan, F.; Mahdavi, H.; Jamshidi, E.; Esmaily, H. Evaluating the Effectiveness of Probiotics in Relieving Constipation in Parkinson’s Disease: A Systematic Review and Meta-Analysis. Heliyon 2023, 9, e14312. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Ji, J.-L.; Li, S.; Cao, X.-P.; Xu, W.; Tan, L.; Tan, C.-C. Efficacy and Safety of Probiotics for the Treatment of Alzheimer’s Disease, Mild Cognitive Impairment, and Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2022, 14, 730036. [Google Scholar] [CrossRef] [PubMed]
- Kuai, X.-Y.; Yao, X.-H.; Xu, L.-J.; Zhou, Y.-Q.; Zhang, L.-P.; Liu, Y.; Pei, S.-F.; Zhou, C.-L. Evaluation of Fecal Microbiota Transplantation in Parkinson’s Disease Patients with Constipation. Microb. Cell Factories 2021, 20, 98. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L.; Suescun, J.; Jiang, Z.-D.; Brown, E.L.; Essigmann, H.T.; Alexander, A.S.; DuPont, A.W.; Iqbal, T.; Utay, N.S.; Newmark, M.; et al. Fecal Microbiota Transplantation in Parkinson’s Disease-A Randomized Repeat-Dose, Placebo-Controlled Clinical Pilot Study. Front. Neurol. 2023, 14, 1104759. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T. Dysfunction of the Microbiota-Gut-Brain Axis in Neurodegenerative Disease: The Promise of Therapeutic Modulation With Prebiotics, Medicinal Herbs, Probiotics, and Synbiotics. J. Evid.-Based Integr. Med. 2020, 25, 2515690X20957225. [Google Scholar] [CrossRef]
- Arora, K.; Green, M.; Prakash, S. The Microbiome and Alzheimer’s Disease: Potential and Limitations of Prebiotic, Synbiotic, and Probiotic Formulations. Front. Bioeng. Biotechnol. 2020, 8, 537847. [Google Scholar] [CrossRef]
- Lye, H.S.; Lee, Y.T.; Ooi, S.Y.; Teh, L.K.; Lim, L.N.; Wei, L.K. Modifying Progression of Aging and Reducing the Risk of Neurodegenerative Diseases by Probiotics and Synbiotics. Front. Biosci. Elite Ed. 2018, 10, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s Risk of Bias Tool for Animal Studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Panaitescu, P.-Ș.; Razniceanu, V.; Mocrei-Rebrean, S.-M.; Filip, A. Signaling and Inflammatory Pathways in Gut Microbiome-Targeted Interventions in Parkinson’s Disease Animal Models—A Systematic Review. Available online: https://www.crd.york.ac.uk/prospero/display_record.php?RecordID=461495 (accessed on 1 April 2024).
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—A Web and Mobile App for Systematic Reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.; McKenzie, J.E.; Sowden, A.; Katikireddi, S.V.; Brennan, S.E.; Ellis, S.; Hartmann-Boyce, J.; Ryan, R.; Shepperd, S.; Thomas, J.; et al. Synthesis without Meta-Analysis (SWiM) in Systematic Reviews: Reporting Guideline. BMJ 2020, 368, l6890. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Meng, J.; Qin, Z.; Yu, Y.; Liang, Y.; Wang, Y.; Min, D. Dysbiosis of Gut Microbiota Inhibits NMNAT2 to Promote Neurobehavioral Deficits and Oxidative Stress Response in the 6-OHDA-Lesioned Rat Model of Parkinson’s Disease. J. Neuroinflamm. 2023, 20, 117. [Google Scholar] [CrossRef]
- Sun, M.-F.; Zhu, Y.-L.; Zhou, Z.-L.; Jia, X.-B.; Xu, Y.-D.; Yang, Q.; Cui, C.; Shen, Y.-Q. Neuroprotective Effects of Fecal Microbiota Transplantation on MPTP-Induced Parkinson’s Disease Mice: Gut Microbiota, Glial Reaction and TLR4/TNF-α Signaling Pathway. Brain. Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Chen, W.; Gao, H.; Che, N.; Xu, M.; Yang, L.; Zhang, Y.; Ye, M. Fecal Microbiota Transplantation Exerts a Protective Role in MPTP-Induced Parkinson’s Disease via the TLR4/PI3K/AKT/NF-κB Pathway Stimulated by α-Synuclein. Neurochem. Res. 2021, 46, 3050–3058. [Google Scholar] [CrossRef]
- Qiao, C.-M.; Zhou, Y.; Quan, W.; Ma, X.-Y.; Zhao, L.-P.; Shi, Y.; Hong, H.; Wu, J.; Niu, G.-Y.; Chen, Y.-N.; et al. Fecal Microbiota Transplantation from Aged Mice Render Recipient Mice Resistant to MPTP-Induced Nigrostriatal Degeneration Via a Neurogenesis-Dependent but Inflammation-Independent Manner. Neurother. J. Am. Soc. Exp. Neurother. 2023, 20, 1405–1426. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, M.; Luo, Y.; Jin, D.; Guo, X.; Yang, W.; Zheng, J.; Zhang, H.; Zhang, L.; Deng, C.; et al. Healthy Human Fecal Microbiota Transplantation into Mice Attenuates MPTP-Induced Neurotoxicity via AMPK/SOD2 Pathway. Aging Dis. 2023, 14, 2193–2214. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, T.; Chen, X.; Zhao, Z.; Chen, Z. Gut Microbiota Relieves Inflammation in the Substantia Nigra of Chronic Parkinson’s Disease by Protecting the Function of Dopamine Neurons. Exp. Ther. Med. 2022, 23, 52. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Ryu, Y.-K.; Chang, D.-H.; Park, H.-Y.; Go, J.; Maeng, S.-Y.; Hwang, D.Y.; Kim, B.-C.; Lee, C.-H.; Kim, K.-S. Agathobaculum Butyriciproducens Shows Neuroprotective Effects in a 6-OHDA-Induced Mouse Model of Parkinson’s Disease. J. Microbiol. Biotechnol. 2022, 32, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Castelli, V.; d’Angelo, M.; Lombardi, F.; Alfonsetti, M.; Antonosante, A.; Catanesi, M.; Benedetti, E.; Palumbo, P.; Cifone, M.G.; Giordano, A.; et al. Effects of the Probiotic Formulation SLAB51 in in Vitro and in Vivo Parkinson’s Disease Models. Aging 2020, 12, 4641–4659. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Carbonell, S.G.; Vásquez-Celaya, L.; García-López, D.; Granados-Patrón, D.; García-Miss, M.D.R.; Álvarez-Cervera, F.J.; Mut-Martín, M.; Parra, I.; Mendieta, L.; Salgado, H.; et al. Chronic Treatment with the Probiotics Lacticaseibacillus Rhamnosus GG and Bifidobacterium Lactis BB12 Attenuates Motor Impairment, Striatal Microglial Activation, and Dopaminergic Loss in Rats with 6-Hydroxydopamine-Induced Hemiparkinsonism. Neuroscience 2022, 507, 79–98. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.-F.; Cheng, Y.-F.; You, S.-T.; Kuo, W.-C.; Huang, C.-W.; Chiou, J.-J.; Hsu, C.-C.; Hsieh-Li, H.-M.; Wang, S.; Tsai, Y.-C. Lactobacillus Plantarum PS128 Alleviates Neurodegenerative Progression in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Mouse Models of Parkinson’s Disease. Brain. Behav. Immun. 2020, 90, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Perez Visñuk, D.; Savoy de Giori, G.; LeBlanc, J.G.; de Moreno de LeBlanc, A. Neuroprotective Effects Associated with Immune Modulation by Selected Lactic Acid Bacteria in a Parkinson’s Disease Model. Nutrition 2020, 79–80, 110995. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chu, C.; Yu, L.; Zhai, Q.; Wang, S.; Zhao, J.; Zhang, H.; Chen, W.; Tian, F. Neuroprotective Effects of Bifidobacterium Breve CCFM1067 in MPTP-Induced Mouse Models of Parkinson’s Disease. Nutrients 2022, 14, 4678. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Wei, J.; Chen, W.; Hong, D.; Chen, T.; Fang, X. Neurotrophic Role of the Next-Generation Probiotic Strain L. lactis MG1363-pMG36e-GLP-1 on Parkinson’s Disease via Inhibiting Ferroptosis. Nutrients 2022, 14, 4886. [Google Scholar] [CrossRef]
- Sun, J.; Li, H.; Jin, Y.; Yu, J.; Mao, S.; Su, K.-P.; Ling, Z.; Liu, J. Probiotic Clostridium Butyricum Ameliorated Motor Deficits in a Mouse Model of Parkinson’s Disease via Gut Microbiota-GLP-1 Pathway. Brain. Behav. Immun. 2021, 91, 703–715. [Google Scholar] [CrossRef]
- Pan, S.; Wei, H.; Yuan, S.; Kong, Y.; Yang, H.; Zhang, Y.; Cui, X.; Chen, W.; Liu, J.; Zhang, Y. Probiotic Pediococcus Pentosaceus Ameliorates MPTP-Induced Oxidative Stress via Regulating the Gut Microbiota-Gut-Brain Axis. Front. Cell. Infect. Microbiol. 2022, 12, 1022879. [Google Scholar] [CrossRef]
- Fang, X.; Zhou, X.; Miao, Y.; Han, Y.; Wei, J.; Chen, T. Therapeutic Effect of GLP-1 Engineered Strain on Mice Model of Alzheimer’s Disease and Parkinson’s Disease. AMB Express 2020, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Srivastav, S.; Neupane, S.; Bhurtel, S.; Katila, N.; Maharjan, S.; Choi, H.; Hong, J.T.; Choi, D.-Y. Probiotics Mixture Increases Butyrate, and Subsequently Rescues the Nigral Dopaminergic Neurons from MPTP and Rotenone-Induced Neurotoxicity. J. Nutr. Biochem. 2019, 69, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Z.; Cheng, S.-H.; Chang, M.-Y.; Lin, Y.-F.; Wu, C.-C.; Tsai, Y.-C. Neuroprotective Effects of Lactobacillus Plantarum PS128 in a Mouse Model of Parkinson’s Disease: The Role of Gut Microbiota and MicroRNAs. Int. J. Mol. Sci. 2023, 24, 6794. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Yu, L.; Li, Y.; Guo, H.; Zhai, Q.; Chen, W.; Tian, F. Lactobacillus Plantarum CCFM405 against Rotenone-Induced Parkinson’s Disease Mice via Regulating Gut Microbiota and Branched-Chain Amino Acids Biosynthesis. Nutrients 2023, 15, 1737. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-H.; Kuo, C.-W.; Hsieh, K.-H.; Shieh, M.-J.; Peng, C.-W.; Chen, Y.-C.; Chang, Y.-L.; Huang, Y.-Z.; Chen, C.-C.; Chang, P.-K.; et al. Probiotics Alleviate the Progressive Deterioration of Motor Functions in a Mouse Model of Parkinson’s Disease. Brain Sci. 2020, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, Z.; Chaiquin, M.; Landrigan, J.; Ayoub, K.; Shail, P.; Rocha, J.; Childers, C.L.; Storey, K.B.; Philpott, D.J.; Sun, H.; et al. The Impact of Dextran Sodium Sulphate and Probiotic Pre-Treatment in a Murine Model of Parkinson’s Disease. J. Neuroinflamm. 2021, 18, 20. [Google Scholar] [CrossRef] [PubMed]
- Parra, I.; Martínez, I.; Vásquez-Celaya, L.; Gongora-Alfaro, J.L.; Tizabi, Y.; Mendieta, L. Neuroprotective and Immunomodulatory Effects of Probiotics in a Rat Model of Parkinson’s Disease. Neurotox. Res. 2023, 41, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.-P.; Nurrahma, B.A.; Kumar, R.; Wu, C.-H.; Yeh, T.-H.; Chiu, C.-C.; Lee, Y.-P.; Liao, Y.-C.; Huang, C.-H.; Yeh, Y.-T.; et al. Probiotic Enhancement of Antioxidant Capacity and Alterations of Gut Microbiota Composition in 6-Hydroxydopamin-Induced Parkinson’s Disease Rats. Antioxidants 2021, 10, 1823. [Google Scholar] [CrossRef]
- Liu, X.; Du, Z.R.; Wang, X.; Sun, X.R.; Zhao, Q.; Zhao, F.; Wong, W.T.; Wong, K.H.; Dong, X.-L. Polymannuronic Acid Prebiotic plus Lacticaseibacillus Rhamnosus GG Probiotic as a Novel Synbiotic Promoted Their Separate Neuroprotection against Parkinson’s Disease. Food Res. Int. Ott. Ont. 2022, 155, 111067. [Google Scholar] [CrossRef]
- Pu, Y.; Chang, L.; Qu, Y.; Wang, S.; Zhang, K.; Hashimoto, K. Antibiotic-Induced Microbiome Depletion Protects against MPTP-Induced Dopaminergic Neurotoxicity in the Brain. Aging 2019, 11, 6915–6929. [Google Scholar] [CrossRef]
- Cui, C.; Hong, H.; Shi, Y.; Zhou, Y.; Qiao, C.-M.; Zhao, W.-J.; Zhao, L.-P.; Wu, J.; Quan, W.; Niu, G.-Y.; et al. Vancomycin Pretreatment on MPTP-Induced Parkinson’s Disease Mice Exerts Neuroprotection by Suppressing Inflammation Both in Brain and Gut. J. Neuroimmune Pharmacol. Off. J. Soc. Neuroimmune Pharmacol. 2023, 18, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Sancandi, M.; De Caro, C.; Cypaite, N.; Marascio, N.; Avagliano, C.; De Marco, C.; Russo, E.; Constanti, A.; Mercer, A. Effects of a Probiotic Suspension SymproveTM on a Rat Early-Stage Parkinson’s Disease Model. Front. Aging Neurosci. 2022, 14, 986127. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kang, L.; Yu, Y.; Long, Y.; Jeffery, J.J.; Cai, W.; Wang, X. Study of Long-Term Biocompatibility and Bio-Safety of Implantable Nanogenerators. Nano Energy 2018, 51, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Robak, L.A.; Yu, M.; Cykowski, M.; Shulman, J.M. Genetics and Pathogenesis of Parkinson’s Syndrome. Annu. Rev. Pathol. 2023, 18, 95–121. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.M.J.; Moreira, P.I.; Ambrósio, A.F.; Alves, C.H. PINK1/PARKIN Signalling in Neurodegeneration and Neuroinflammation. Acta Neuropathol. Commun. 2020, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Malpartida, A.B.; Williamson, M.; Narendra, D.P.; Wade-Martins, R.; Ryan, B.J. Mitochondrial Dysfunction and Mitophagy in Parkinson’s Disease: From Mechanism to Therapy. Trends Biochem. Sci. 2021, 46, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, E.; Pointer, C.B.; Klegeris, A. The Role of Mitochondrial Damage-Associated Molecular Patterns in Chronic Neuroinflammation. Mediators Inflamm. 2019, 2019, 4050796. [Google Scholar] [CrossRef]
- Borsche, M.; Pereira, S.L.; Klein, C.; Grünewald, A. Mitochondria and Parkinson’s Disease: Clinical, Molecular, and Translational Aspects. J. Park. Dis. 2021, 11, 45–60. [Google Scholar] [CrossRef]
- Tansey, M.G.; Wallings, R.L.; Houser, M.C.; Herrick, M.K.; Keating, C.E.; Joers, V. Inflammation and Immune Dysfunction in Parkinson Disease. Nat. Rev. Immunol. 2022, 22, 657–673. [Google Scholar] [CrossRef]
- Konnova, E.A.; Swanberg, M. Animal Models of Parkinson’s Disease. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; Stoker, T.B., Greenland, J.C., Eds.; Codon Publications: Brisbane, Australia, 2018; ISBN 978-0-9944381-6-4. [Google Scholar]
- Deng, I.; Corrigan, F.; Zhai, G.; Zhou, X.-F.; Bobrovskaya, L. Lipopolysaccharide Animal Models of Parkinson’s Disease: Recent Progress and Relevance to Clinical Disease. Brain Behav. Immun.-Health 2020, 4, 100060. [Google Scholar] [CrossRef]
- Beckstead, M.J.; Howell, R.D. Progressive Parkinsonism Due to Mitochondrial Impairment: Lessons from the MitoPark Mouse Model. Exp. Neurol. 2021, 341, 113707. [Google Scholar] [CrossRef]
- Lin, C.-H.; Lai, H.-C.; Wu, M.-S. Gut-Oriented Disease Modifying Therapy for Parkinson’s Disease. J. Formos. Med. Assoc. Taiwan Yi Zhi 2023, 122, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. Toll-like Receptors in the Pathogenesis of Neuroinflammation. J. Neuroimmunol. 2019, 332, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Heidari, A.; Yazdanpanah, N.; Rezaei, N. The Role of Toll-like Receptors and Neuroinflammation in Parkinson’s Disease. J. Neuroinflamm. 2022, 19, 135. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The Nuclear Factor NF-kappaB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, T.G. Role of Nuclear Factor Kappa B (NF-κB) Signalling in Neurodegenerative Diseases: An Mechanistic Approach. Curr. Neuropharmacol. 2020, 18, 918–935. [Google Scholar] [CrossRef] [PubMed]
- Dolatshahi, M.; Ranjbar Hameghavandi, M.H.; Sabahi, M.; Rostamkhani, S. Nuclear Factor-Kappa B (NF-κB) in Pathophysiology of Parkinson Disease: Diverse Patterns and Mechanisms Contributing to Neurodegeneration. Eur. J. Neurosci. 2021, 54, 4101–4123. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB Pathway for the Therapy of Diseases: Mechanism and Clinical Study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Gordon, R.; Albornoz, E.A.; Christie, D.C.; Langley, M.R.; Kumar, V.; Mantovani, S.; Robertson, A.A.B.; Butler, M.S.; Rowe, D.B.; O’Neill, L.A.; et al. Inflammasome Inhibition Prevents α-Synuclein Pathology and Dopaminergic Neurodegeneration in Mice. Sci. Transl. Med. 2018, 10, eaah4066. [Google Scholar] [CrossRef]
- Li, Y.; Xia, Y.; Yin, S.; Wan, F.; Hu, J.; Kou, L.; Sun, Y.; Wu, J.; Zhou, Q.; Huang, J.; et al. Targeting Microglial α-Synuclein/TLRs/NF-kappaB/NLRP3 Inflammasome Axis in Parkinson’s Disease. Front. Immunol. 2021, 12, 719807. [Google Scholar] [CrossRef]
- Trudler, D.; Nazor, K.L.; Eisele, Y.S.; Grabauskas, T.; Dolatabadi, N.; Parker, J.; Sultan, A.; Zhong, Z.; Goodwin, M.S.; Levites, Y.; et al. Soluble α-Synuclein-Antibody Complexes Activate the NLRP3 Inflammasome in hiPSC-Derived Microglia. Proc. Natl. Acad. Sci. USA 2021, 118, e2025847118. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.N.; Nguyen, H.D.; Kim, Y.J.; Nguyen, T.T.; Lai, T.T.; Lee, Y.K.; Ma, H.-I.; Kim, Y.E. Role of NLRP3 Inflammasome in Parkinson’s Disease and Therapeutic Considerations. J. Park. Dis. 2022, 12, 2117–2133. [Google Scholar] [CrossRef] [PubMed]
- Nakano, N.; Matsuda, S.; Ichimura, M.; Minami, A.; Ogino, M.; Murai, T.; Kitagishi, Y. PI3K/AKT Signaling Mediated by G Protein-coupled Receptors Is Involved in Neurodegenerative Parkinson’s Disease (Review). Int. J. Mol. Med. 2017, 39, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.K.; Jha, N.K.; Kar, R.; Ambasta, R.K.; Kumar, P. P38 MAPK and PI3K/AKT Signalling Cascades inParkinson’s Disease. Int. J. Mol. Cell. Med. 2015, 4, 67–86. [Google Scholar] [PubMed]
- Golpich, M.; Amini, E.; Hemmati, F.; Ibrahim, N.M.; Rahmani, B.; Mohamed, Z.; Raymond, A.A.; Dargahi, L.; Ghasemi, R.; Ahmadiani, A. Glycogen Synthase Kinase-3 Beta (GSK-3β) Signaling: Implications for Parkinson’s Disease. Pharmacol. Res. 2015, 97, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Agrawal, A.; Verma, A.; Dubey, N. The PI3K-AKT Pathway: A Plausible Therapeutic Target in Parkinson’s Disease. Exp. Mol. Pathol. 2023, 129, 104846. [Google Scholar] [CrossRef] [PubMed]
- Fock, E.; Parnova, R. Mechanisms of Blood-Brain Barrier Protection by Microbiota-Derived Short-Chain Fatty Acids. Cells 2023, 12, 657. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Sheng, S.; Zhang, F. Relationship Between Gut Bacteria and Levodopa Metabolism. Curr. Neuropharmacol. 2023, 21, 1536–1547. [Google Scholar] [CrossRef]
- Zhong, Z.; Ye, M.; Yan, F. A Review of Studies on Gut Microbiota and Levodopa Metabolism. Front. Neurol. 2023, 14, 1046910. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Quagliariello, A.; Schiavoni, M.; Petito, V.; Russo, A.; Reddel, S.; Del Chierico, F.; Ianiro, G.; Scaldaferri, F.; Neri, M.; et al. Towards a Disease-Associated Common Trait of Gut Microbiota Dysbiosis: The Pivotal Role of Akkermansia Muciniphila. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2020, 52, 1002–1010. [Google Scholar] [CrossRef]
- Ouyang, J.; Lin, J.; Isnard, S.; Fombuena, B.; Peng, X.; Marette, A.; Routy, B.; Messaoudene, M.; Chen, Y.; Routy, J.-P. The Bacterium Akkermansia Muciniphila: A Sentinel for Gut Permeability and Its Relevance to HIV-Related Inflammation. Front. Immunol. 2020, 11, 645. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Choi, Y.; Kim, D.-K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.-S.; Jee, Y.-K.; Gho, Y.S.; et al. Akkermansia Muciniphila-Derived Extracellular Vesicles Influence Gut Permeability through the Regulation of Tight Junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, J.; Young, L.H.; Caplan, M.J. AMP-Activated Protein Kinase Regulates the Assembly of Epithelial Tight Junctions. Proc. Natl. Acad. Sci. USA 2006, 103, 17272–17277. [Google Scholar] [CrossRef]
- Ghotaslou, R.; Nabizadeh, E.; Memar, M.Y.; Law, W.M.H.; Ozma, M.A.; Abdi, M.; Yekani, M.; Kadkhoda, H.; Hosseinpour, R.; Bafadam, S.; et al. The Metabolic, Protective, and Immune Functions of Akkermansia Muciniphila. Microbiol. Res. 2023, 266, 127245. [Google Scholar] [CrossRef]
- Ottman, N.; Reunanen, J.; Meijerink, M.; Pietilä, T.E.; Kainulainen, V.; Klievink, J.; Huuskonen, L.; Aalvink, S.; Skurnik, M.; Boeren, S.; et al. Pili-like Proteins of Akkermansia Muciniphila Modulate Host Immune Responses and Gut Barrier Function. PLoS ONE 2017, 12, e0173004. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Saier, M.H. Gut Bacteroides Species in Health and Disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef]
- Bi, M.; Liu, C.; Wang, Y.; Liu, S.-J. Therapeutic Prospect of New Probiotics in Neurodegenerative Diseases. Microorganisms 2023, 11, 1527. [Google Scholar] [CrossRef]
- Shao, X.; Sun, S.; Zhou, Y.; Wang, H.; Yu, Y.; Hu, T.; Yao, Y.; Zhou, C. Bacteroides Fragilis Restricts Colitis-Associated Cancer via Negative Regulation of the NLRP3 Axis. Cancer Lett. 2021, 523, 170–181. [Google Scholar] [CrossRef]
- King, C.H.; Desai, H.; Sylvetsky, A.C.; LoTempio, J.; Ayanyan, S.; Carrie, J.; Crandall, K.A.; Fochtman, B.C.; Gasparyan, L.; Gulzar, N.; et al. Baseline Human Gut Microbiota Profile in Healthy People and Standard Reporting Template. PLoS ONE 2019, 14, e0206484. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Luo, L.; Liang, W.; Yin, Q.; Guo, J.; Rush, A.M.; Lv, Z.; Liang, Q.; Fischbach, M.A.; Sonnenburg, J.L.; et al. Bifidobacterium Alters the Gut Microbiota and Modulates the Functional Metabolism of T Regulatory Cells in the Context of Immune Checkpoint Blockade. Proc. Natl. Acad. Sci. USA 2020, 117, 27509–27515. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fang, D.; Wang, Z.; Liu, Y. Sleep Deprivation and Gut Microbiota Dysbiosis: Current Understandings and Implications. Int. J. Mol. Sci. 2023, 24, 9603. [Google Scholar] [CrossRef]
- Neculicioiu, V.S.; Colosi, I.A.; Costache, C.; Sevastre-Berghian, A.; Clichici, S. Time to Sleep?—A Review of the Impact of the COVID-19 Pandemic on Sleep and Mental Health. Int. J. Environ. Res. Public Health 2022, 19, 3497. [Google Scholar] [CrossRef] [PubMed]
- Rethlefsen, M.L.; Kirtley, S.; Waffenschmidt, S.; Ayala, A.P.; Moher, D.; Page, M.J.; Koffel, J.B.; PRISMA-S Group. PRISMA-S: An extension to the PRISMA Statement for Reporting Literature Searches in Systematic Reviews. Syst. Rev. 2021, 10, 39. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
| In vivo murine Parkinson’s disease models | In vitro, in silico, in vivo human or non-murine models, all review articles |
| Intervention—Fecal microbiota transplantation, Antibiotic, Prebiotic, Probiotic, or Synbiotic supplementation | Dietary, nutraceutical, fatty acids, and fatty acid derivate interventions. |
| Outcomes: dopaminergic cell loss; at least one of the others: cytokines, neuroinflammatory markers, or motor behavior tests | High risk of bias |
| Data Category | Outcomes | Extracted data |
|---|---|---|
| Study design | Parkinson’s induction model | Animals and induction substance |
| Dose, time, frequency, and route of administration | ||
| Other data (vehicle, anesthetics, attrition) | ||
| Intervention | Type of intervention—Probiotic strain, FMT donor, Antibiotics | |
| Dose, time, frequency, and route of administration | ||
| Other data (vehicle, anesthetics, attrition) | ||
| Primary outcomes | Dopaminergic cell loss | Tyrosine Hydroxilase (TH) |
| Microglial activation | Ionized calcium-binding adaptor molecule 1 (Iba1) | |
| Astrocyte activation | Glial fibrillary acidic protein (GFAP) | |
| Cytokines | IL-6, IL-1β, TNF-α | |
| Motor behavioral tests | Rotarod test, Narrow beam test, Pole test, Open field test | |
| Traction test, Grip strength, Hanging test, Cylinder test | ||
| Apomorphine-induced/Amphetamine-induced rotation test | ||
| Gait analysis, Elevated body swing test, Sticker test | ||
| Microbiome analysis | Relative abundance analysis | |
| Secondary outcomes | Cytokines | IL-10, IL-1α |
| Neuroinflammation markers and signaling pathways | BDNF, Glial cell line-derived neurotrophic factor (GDNF), Proliferator-activated receptor γ (PPaR γ), Cyclooxygenase-2 (COX2), iNOS, PI3K, Protein kinase B (Akt), NF-κB | |
| Microbiome analysis | Alpha and beta diversity | |
| Other outcomes | α-syn, LPS, ZO-1, Occludin, Claudin, SCAF |
| Study | Probiotic Strains |
|---|---|
| Castelli et al. [84] | S. thermophilus, B. longum, B. breve, B. infantis, L. acidophilus, L. plantarum, L. paracasei, L. Delbrueckii subsp. bulgaricus, and L. brevis |
| Cuevas-Carbonell et al. [85] | L. rhamnosus GG, B. animalis subsp. lactis BB-12 |
| Dwyer et al. [97] | L. plantarum, L. delbrueckii subsp. bulgaricus, L. paracasei, L. acidophilus, B. breve, B. longum, B. infantis, and S. salivarius subsp. thermophilus |
| Hsieh et al. [96] | B. bifidum, B. longum, L. rhamnosis, L. rhamnosus GG, L. plantarum LP28, and L. lactis subsp. lactis |
| Parra et al. [98] | L. rhamnosus GG, B. animalis subsp. lactis BB-12 |
| Perez Visnuk el al [87] | L. plantarum CRL 2130, S. thermophilus CRL 808, and S. thermophilus CRL 807 |
| Srivastav et al. [93] | L. rhamnosus GG, B. animalis subsp. lactis BB-12, and L. acidophilus LA-5 |
| Study, Year | PD Model | Interv. | TH | Iba-1 | GFAP | IL-6 | TNF-alfa | IL-1beta | Behavioural Test Performance | Gut Microbiome Relative Abundance |
|---|---|---|---|---|---|---|---|---|---|---|
| Yu et al., 2023 [77] | 6-OHDA, rat | FMT | SN: ↑ * | RR: ↑ ***; APO: ↑ ***; OF: ↑ ** | ||||||
| Sun et al., 2018 [78] | MPTP, mice | FMT | SN ↑ **; STR ↑ * | SN ↓ *** | SN ↓ *** | STR ↓ **; Colon ↓ ** | PT: ↑ *** | ↑ Clostridiales * ↓ Proteobacteria **; Turicibacterales *, Enterobacterales ** | ||
| Zhong et al., 2021 [79] | MPTP, mice | FMT | SN IHC ↑ **; WB ↑ *** | SN ↓ **; STR ↓ * | TT: ↑ ***; PT: ↑ ** | |||||
| Qiao et al., 2023 [80] | MPTP, mice | FMT | AM + MPTP IF ↑ ##, WB ns. | ns | SN ns; Colon ns | SN ns; Colon ns | SN ns; colon ns | RR: AM + MPTP ↓ #, TT ns; PT: AM + MPTP ↓ ## | AM—↑ Lactobacillus ##; ↑ Proteobacteria #, ↑ Desulfovibrionales # ↑ Dubosiella #, ↑ Helicobacter #, ↑ Campylobacter #, ↑ Odoribacter #, ↑ Parvibacter #, ↑ Anaeroplasma # ↓ AM—Ruminococcacea UCG-014 ##, ↓ Eubacterium xylanophilum group # | |
| Xie et al., 2023 [81] | MPTP, mice | FMT | STR PD ↓ *, HC ↑ ***; SNpc PD↓ *** HC↑ *** | SNpc PD ↑ ***, HC ↓ **; STR PD ↑ *** HC ↓ **; Colon PD ↑ ***, HC ↓ ** | SNpc PD ↑ *** HC ↓ ***; STR PD ↑ ***; HC ↓ *** | Colon: PD ↑ **, HC ↓ * | RR: PD ↓ **; HC ↑ ***; PT: PD ↓ ***; HC ↑ ***. | PD: ↑ Verrucomicrobiota *** ↑ Akkermansia *** ↓ Eubacterium xylanophilum group **, ↓ Lachnospiraceae unclassified * HC: ↓ Desulfobacterota **, Desulfovibrio ** | ||
| Zhang et al., 2021 [82] | MPTP + probenecid, mice | FMT | SN ↑ * | SN M1 ↓ *** | SN ↓ ** | SN ↓ *, Colon: ↓ * | RR: ↑ *; PT: ↑ *; OF: ↑ ** | ↑ Blautia * ↓ Proteobacteria **, ↓ Tenericutes *; ↓ Anaerostipes **, ↓ Bifidobacterium *, ↓ ASF356 **, ↓ Ruminococcus ** | ||
| Zhao et al., 2021 [47] | Rotenone, mice | FMT | SN: IF ↑ ***, Mid-brain: WB ↑ * | SN ↓ *** | SN ↓ *** | Mid-brain ↓ ***; SN ↓ *; Colon: ↓ *** | Mid-brain: ↓ ***; SN ↓ **; Colon: ↓ *** | Mid-brain: ↓ ***; SN ↓ *; Colon: ↓ *** | RR: ↑ ***; GT: ↑ **; PT: ↑ **; ST: ↑ ***. | ↑ Proteobacteria **, ↑ Helicobacteraceae ***, ↑ Enterobacteriaceae *, ↑ Lactobacillaceae **; ↑ Barnesiella *, ↑ Butyricicoccus **, ↑ Helicobacter ***, ↑ Roseburia ** ↓ Verrucomicrobia *** ↓ Akkermansia ***, ↓ Coriobacteriaceae **, ↓ Desulfovibrio * |
| Lee et al., 2022 [83] | 6-OHDA, mice | PRO | ns | ns | SN ↓ ** | AMPH: ↑ * | ||||
| Castelli et al., 2020 [84] | 6-OHDA, mice | PRO | STR ↑ *; SN ↑ ** | STR ↓ ** | STR ↓ * | APO: ↑ **; EBST: ↑ *** | ||||
| Cuevas-Carbonell et al., 2022 [85] | 6-OHDA, rats | PRO | STR ↑ ***; SNpc ↑ * | STR ↓ ** | BT: ↑ *; APO: ns; CT: ns; GA: ns. | |||||
| Liao et al., 2020 [86] | MPTP, mice | PRO | SN ↑ ***; STR ↑ *** | STR: ↓ ** | STR ↓ * | STR ↓ ** | STR ↓ *** | STR ↓ ** | RR: ↑ ***; PT: ↑ ***; BT: ↑ *** | ↑ Lactobacillus plantarum *** ↓ Enterobacteriaceae *** |
| Perez Visnuk et al., 2020 [87] | MPTP + probenecid, mice | PRO | SNpc Mix ↑ * | Serum Mix ↓ * | Serum Mix ns; Brain: Mix ↓ * | PT: Mix ↑ *, 2130 ↑ *, 807 ↑ *, 808 ↑ *; BT: Mix ↑ *, 2130 ↑ *, 807 ↑ *, 808 ↑ *; ST: Mix ↑ * | ||||
| Li et al., 2022 [88] | MPTP, mice | PRO | SN ↑ ***; STR ↑ *** | STR ↓ *** | STR ↓ *** | STR ↓ ***; Colon ↓ *** | STR ↓ ***; Colon ↓ *** | STR ↓ ***; Colon ↓ * | RR: ↑ **; PT: ↑ ***; OF: ↑ ***; BT: ↑ *** | ↑ Akkermansia *, ↑ Bifidobacterium ** ↓ Bacteroides **, ↓ Escherichia-Shigella *, ↓ Dubosiella **, ↓ Lactobacillus ** |
| Yue et al., 2022 [89] | MPTP, mice | PRO | SN IHC↑ *; WB ↑ ** | HT: ↑ ** PT: ↑ **; OF: GLP-1 ↑ ** | ||||||
| Sun et al., 2021 [90] | MPTP, mice | PRO | SN ↑ * | PT: ↑ **; OF: ↑ **; BT: ↑ ** | ↑ Verrucomicrobia **; ↑ Akkermansia: *, ↓ PrevotellaceaeNK3B31 **; ↓ Alistipes *; ↓ Odoribacter * | |||||
| Pan et al., 2022 [91] | MPTP, mice | PRO | SN ↑ ** | RR: ↑ **; PT: ↑ **; BT: ↑ ** | ↑ Bacteroidota **, ↑ Muribaculaceae **, ↑ Lachnospiraceae *, ↑ Defluviitaleaceae *, ↓ Proteobacteria **, ↓ Firmicutes *, ↓ Erysipelotrichaceae *, ↓ Enterococcaceae **, ↓ Dubosiella *, ↓ Enterococcus ** | |||||
| Fang et al., 2020 [92] | MPTP, mice | PRO | ns | PT: ↑ *; OF: ↑ ** | ns | |||||
| Srivastav et al., 2019 [93] | MPTP, mice | PRO | SN ↑ *, STR ↑ ** | SNpc ↓ ** | SNpc ↓ ** | BT: ↑ *; CBT ↑ ***; GA: ↑ *; CT: ↑ *** | ||||
| Srivastav et al., 2019 [93] | rotenone, mice | PRO | SN ↑ **, STR ↑ ** | SNpc ↓ *** | SNpc ↓ *** | BT: ↑ *; CBT ↑ ***; GA: ↑ ***; CT: ↑ *** | ||||
| Lee et al., 2023 [94] | rotenone, mice | PRO | SN ↑ **; STR ↑ ** | SN ↓ * | SN ↓ *** | RR: ↑ *; Beam: ↑ *** | ↑ Bifidobacterium ***, ↑ Ruminiclostridium 6 ***, ↑ Adlercreutzia ** ↑ ASF356 ** ↑ Acetatifactor * | |||
| Chu et al., 2023 [95] | rotenone, mice | PRO | SN ↑ * | Nigro-striatal ↓ ** | Nigro-striatal ↓ * | Mid-brain: ↓ **; Colon ↓ * | Mid-brain ↓ **; Colon: ↓ ** | Mid-brain ↓ *; Colon: ns | RR: ↑ *; PT: ↑ 50 cm **, 70 cm *; OF: ↑ 5 mm *; ↑ 10 mm ** | ↑ Actinobacteria ***, ↑ Bifidobacterium *, ↑ Faecalibaculum *, ↑ Turicibacter * ↓ Firmicutes *, ↓ Bacteroidetes ***, ↓ Alistipes *, ↓ Akkermansia *, ↓ Bilophila *, ↓ Ruminococcaceae UCG 004 *, ↓ Ruminococcaceae UCG 009 * |
| Hsieh et al., 2020 [96] | MitoPark PD, mice | PRO | SNpc ↑ * | RR: ↑ ***; BT: ↑ ***; GA: ↑ * | ||||||
| Dwyer et al., 2021 [97] | LPS + PQ, mice | PRO | SNpc ↑ ** | ns | ns | ns | ns | ns | RR: -; Micromax: - | ↑ Streptococcaceae *** |
| Parra et al., 2023 [98] | LPS, rats | PRO | ns | SN ns; STR ↓ *; | PT: NS; BT: swing phase ↑ **; CT: ns | |||||
| Tsao et al., 2021 [99] | 6-OHDA, rats | SYM | STR: PRO ↑ *, PRE ↑ * SYN ↑ *; SN: PRO ↑ *, PRE ↑ * SYN ↑ *; | STR PRO ↓ *, PRE ↓ * and SYN ↓ * | RR: ↑ RO *, PRE *, SYM *; APO: ↑ PRO *, PRE * and SYM * | SYN: ↑ Ruminococcaceae *, ↓ Aggregatibacter *, ↓ Balutia *, ↓ Coprococcus *, ↓ Eubacterium *, ↓ Prevotella * PRO: ↑ Ruminococcaceae *, ↓ Propionibacterium *, ↓ Clostridium * PRE: ↑ Prevotella *, ↑ Elizabethkingia *, ↑ Eggerthella *, ↑ Faecalibacterium *, ↑ Mitsuokella *, ↑ Succinatimonas *, ↑ Bifidobacterium *, ↓ Lactobacillus * | ||||
| Liu et al., 2022 [100] | MPTP, mice | SYM | Midbrain: SYN ↑ **; STR: SYN ↑ ***, PRE ↑ *, PRO ↑ * | STR ns | STR PRE ↓ *** | STR ns | GT: SYN ↑ **; OF: PRE *, PRO: * ↑, SYN: ↑ ***; | SYM: ↑ Lactobacillales *, ↑ Lactobacillus *, ↓ Bacteriodaceae **, ↓ Bacteroides ** PRO: ↑ Clostridia, ↑ Ruminococcaceae **, ↓ Lactobabilalles *, ↓ Lactobacillus * PRE: ↓ Lactobabilalles **, ↓ Lactobacillus ** all groups ↓ Turicibacterales, ↓ Turicibacter *** | ||
| Koutzoumis et al., 2019 [41] | 6-OHDA, rat | ATB | STR ↑ **; SN ns | STR ns; | STR: ↓ * | STR: ↓ * | AMPH: ↑ *; GA: ↑ *; CT: ↑ * | ns | ||
| Hong et al., 2022 [42] | MitoPark PD, mice | ATB | ns | SN ↓ * | Serum ↓ * | Serum ↓ * | Serum ↓ * | BT: ↑ **; GA: ↑ * | ↓ Prevotellaceae UCG-001 ** | |
| Pu et al., 2019 [101] | MPTP, mice | ATB | SN, STR ↑ ** | ↑ Bacteroidetes ***, ↑ Proteobacteria ***; ↑ Robinsoniella *, ↑ Dorea *, ↑ Parabacteroides *** ↓ Firmicutes **; ↓ Lactobacillus *** | ||||||
| Cui et al., 2023 [102] | MPTP, mice | ATB | ns | SNpc ↓ *** | SNpc ↓ *** | STR ↓ ***; Colon↓ * | TT: ↑ ***; PT: ↑ *** | ↑ Verrucomicrobia **, ↑ Ileibacterium *, ↑ Akkermansia **, ↑ Blautia * ↓ Actinobacteria *, ↓ Bifidobacteriales *, ↓ Coriobacteriales ***; ↓ Dubosiella *, ↓ Bifidobacterium * |
| Study | Secondary Outcomes |
|---|---|
| Sun et al., 2018 [78] | Alpha div. NS; |
| Zhong et al., 2021 [79] | a-syn ↓ ***, TLR4 ↓ Str ***, SN ***; ↓ p-PI3K/PI3K Str ***, SN **, ↓ p-Akt/Akt Str ***, SN ***, NF-κB ↓ Str *, SNpc ***, ↓ AA ***, PA ***, BA ***, N-VA *** |
| Qiao et al., 2023 [80] | Alpha div. NS; Occludin, Claudin-1 NS, IL-10 Colon NS, SN NS; AM: ↓ BA # IBA ###, IVA ### VA ##, PA ###, AA ##, Colon iNOS NS; |
| Xie et al., 2023 [81] | PD: GDNF ↑ SNpc ***, Str ***; HC: ↓ SNpc ***, Str ***; |
| Zhang et al., 2021 [82] | IL-10 ↑ SN *, Colon *, iNOS SN ↓ * |
| Zhao et al., 2021 [47] | Alpha div.: ↑ Shannon *, ↑ Simpson *; Midbrain a-syn ↓ ***, LPS ↓ SN, Colon ***, Fecal ***, ZO-1 ↑ **, Occludin ↑ ***, Claudin5 ↑ ***, TLR4 ↓ SN ***, Colon ***, COX2 ↓ midbrain *, colon **, SN **; iNOS ↓ midbrain **, colon ***, SN **; NF-κB serum ↓ *** |
| Lee et al., 2022 [83] | ↑ pAKT/AKT ** |
| Castelli et al., 2020 [84] | BDNF ↑ SN ***, Str *; PPaRγ↑ Str ***, SN ***; Pi3K ↑ ***; p-AKT ↑ ***; NF-κB ↓ Str **, SN *** |
| Liao et al., 2020 [86] | Mature BDNF ↑ *; Alpha div.: ↑ Shannon *, Evenness ***, ↓ Chao1 *; ↓ LPS synthesizing module *, SCFA NS |
| Perez Visnuk et al., 2020 [87] | Mixture ↑ IL-10 * |
| Li et al., 2022 [88] | BDNF ↑ Striatum ***; GDNF ↑ Striatum ***; Alpha diversity: ↑ Shannon *, Simpson **; ZO-1 ↓ Striate *, colon *, claudin-1 ↓ Striate *, colon *; ↑ IL-10 colon ***, ↑ AA ***, BA ***, IBA ***, IVA *** |
| Yue et al., 2022 [89] | BDNF ↑ *; GDNF ↑ *; Alpha diversity: Simpson ↑ *; a-syn ↓ aggregation **, ZO-1 ↓ colon *, occludin ↓ colon * |
| Sun et al., 2021 [90] | Alpha diversity NS; TLR4 ↓ Str **, Colon *; NF-κB ↓ Str **, Colon *; ↓ AA ***, PA **, BA *, VA ** |
| Pan et al., 2022 [91] | Alpha diversity: ↑ Shannon **, ↓ Simpson **; α-syn ↓ in SN ** |
| Fang et al., 2020 [92] | TLR4↓ *, Akt NS |
| Srivastav et al., 2019 [93] | MPTP: BDNF ↑ **; GDNF ↑ **; ↑ pPI3K/PI3K *, ↑ pAkt/Akt * |
| Srivastav et al., 2019 [93] | Rotenone: BDNF ↑ *; GDNF ↑ **; |
| Lee et al., 2023 [94] | BDNF NS; Alpha diversity NS; IL-10 ↑ SN ***, iNOS ↓ |
| Chu et al., 2023 [95] | Alpha diversity ↓ *; Occludin ↑ *; ZO-1 * ↑ |
| Tsao et al., 2021 [99] | Synbiotic group: Alpha diversity: Shannon ↑ *; All groups ↑ total acid *, PA *, BA * |
| Liu et al., 2022 [100] | Synbiotic: BDNF ↑ Striatum *; GDNF ↑ Striatum *; Prebiotic: ↑ AA ***, BA *** |
| Koutzoumis et al., 2019 [41] | ↓ COX2 * |
| Hong et al., 2022 [42] | Alpha diversity ↓ *; ↓ serum occludin * and claudin-5 ** |
| Pu et al., 2019 [101] | Alpha diversity: Shannon ↓ **, Chao1 ↓ ***, ACE ↓ ** |
| Cui et al., 2023 [102] | Alpha diversity: Chao1 ↓ ***, Obs. species index ↑ ***; TLR4 ↓ Str *, colon *; NF-κB ↓ Str *, colon * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panaitescu, P.-Ș.; Răzniceanu, V.; Mocrei-Rebrean, Ș.-M.; Neculicioiu, V.S.; Dragoș, H.-M.; Costache, C.; Filip, G.A. The Effect of Gut Microbiota-Targeted Interventions on Neuroinflammation and Motor Function in Parkinson’s Disease Animal Models—A Systematic Review. Curr. Issues Mol. Biol. 2024, 46, 3946-3974. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46050244
Panaitescu P-Ș, Răzniceanu V, Mocrei-Rebrean Ș-M, Neculicioiu VS, Dragoș H-M, Costache C, Filip GA. The Effect of Gut Microbiota-Targeted Interventions on Neuroinflammation and Motor Function in Parkinson’s Disease Animal Models—A Systematic Review. Current Issues in Molecular Biology. 2024; 46(5):3946-3974. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46050244
Chicago/Turabian StylePanaitescu, Paul-Ștefan, Vlad Răzniceanu, Ștefania-Maria Mocrei-Rebrean, Vlad Sever Neculicioiu, Hanna-Maria Dragoș, Carmen Costache, and Gabriela Adriana Filip. 2024. "The Effect of Gut Microbiota-Targeted Interventions on Neuroinflammation and Motor Function in Parkinson’s Disease Animal Models—A Systematic Review" Current Issues in Molecular Biology 46, no. 5: 3946-3974. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb46050244