
17β-Estradiol (E2) Activates Matrix Mineralization through Genomic/Nongenomic Pathways in MC3T3-E1 Cells

 , , , , , , ,
, , , , , , ,
Abstract
:
1. Introduction
2. Results
2.1. E2 Stimulation Triggers Matrix Mineralization of MC3T3-E1 Cells
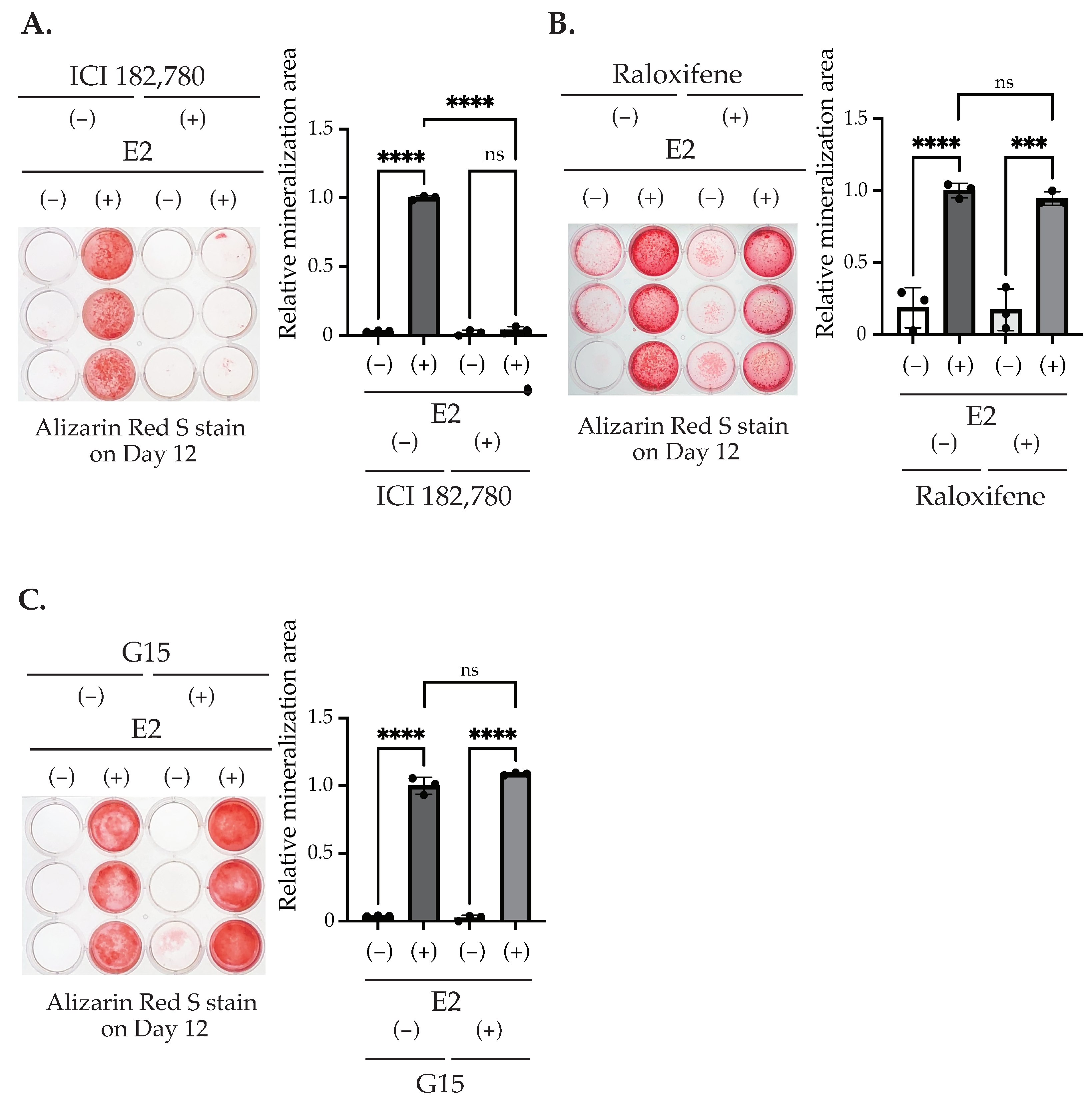
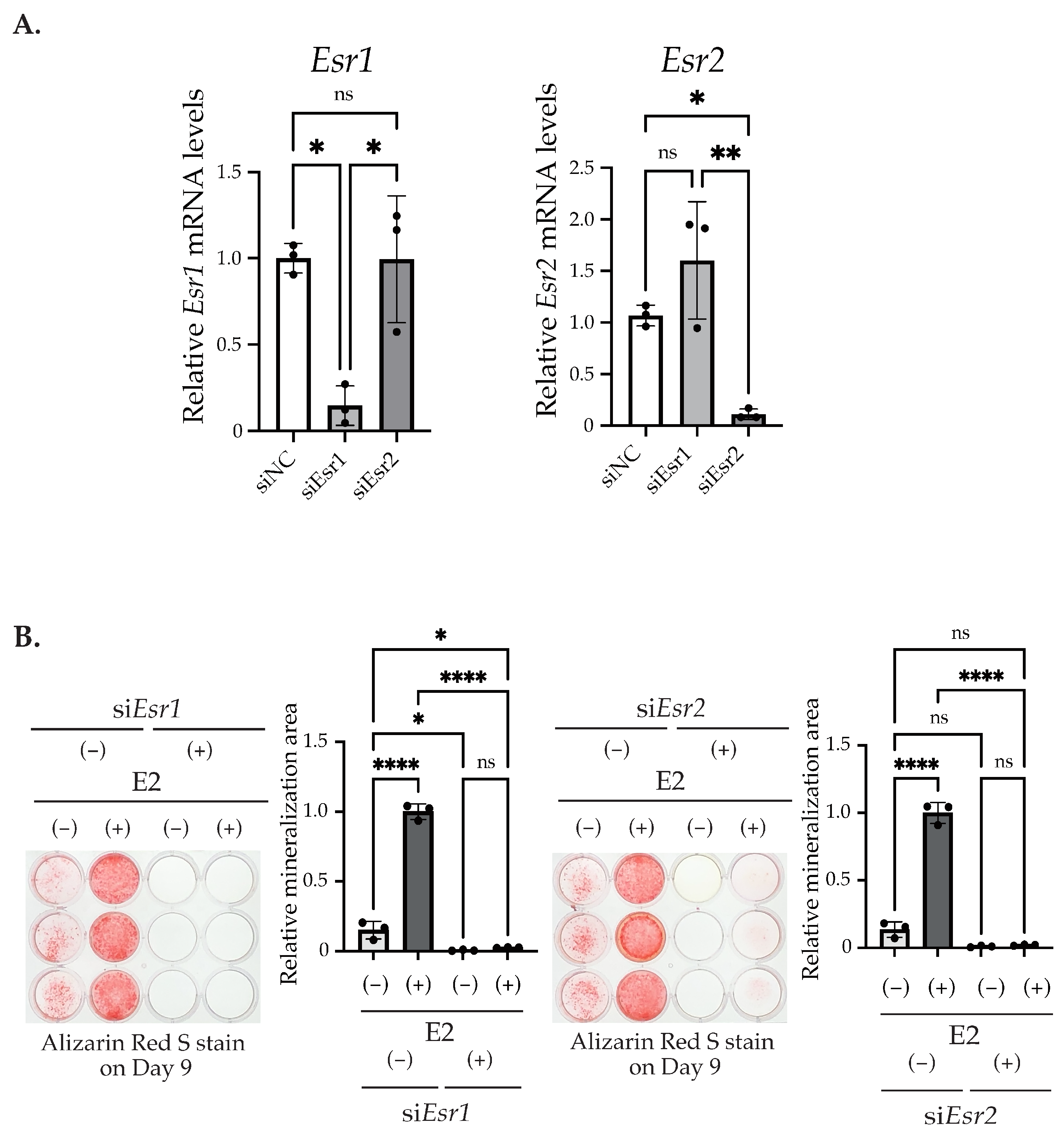
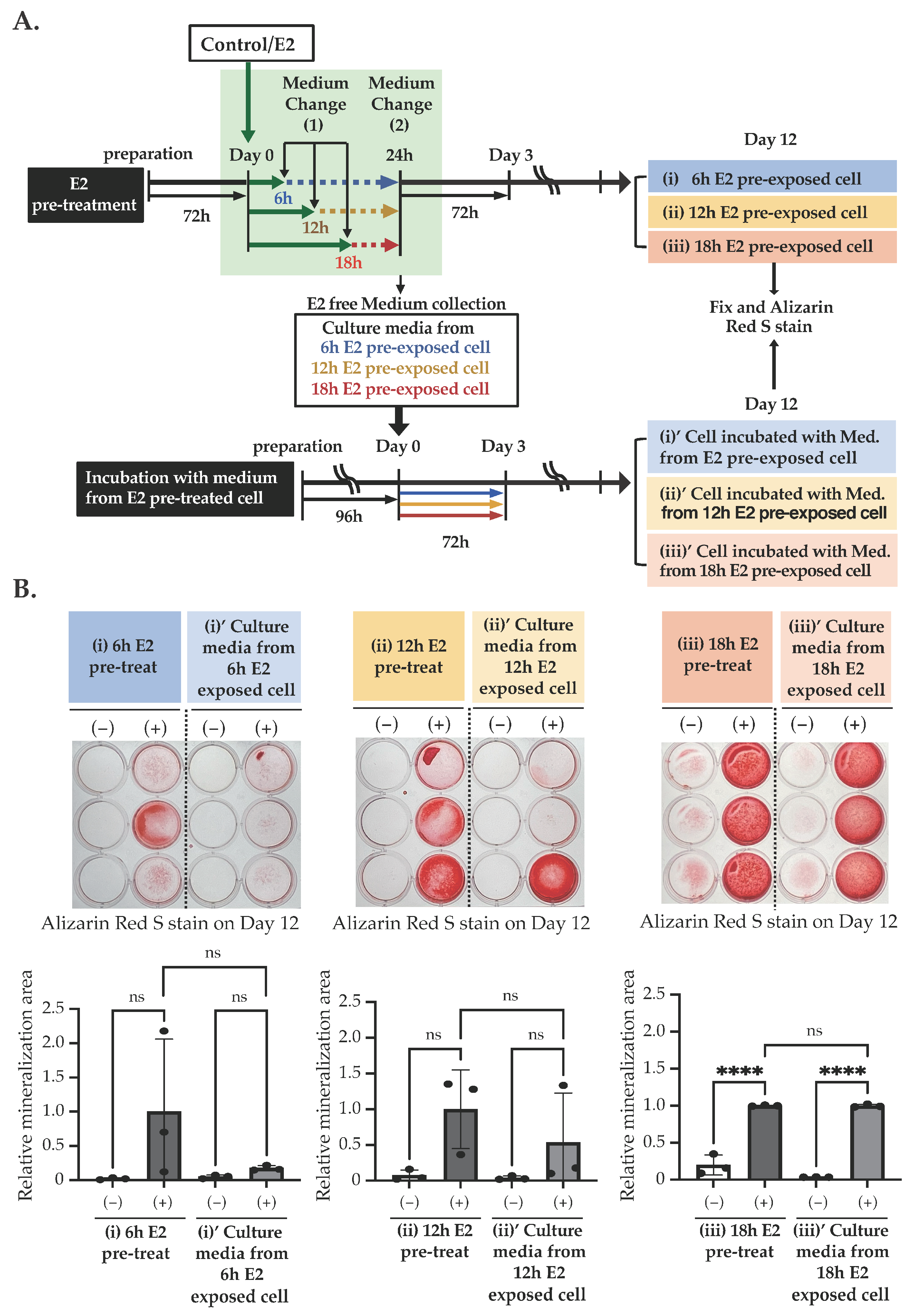
2.2. E2 Stimulation Induces the Matrix Mineralization of MC3T3-E1 via the ER Pathway Independent from Nuclear Translocation and Transcription Regulation
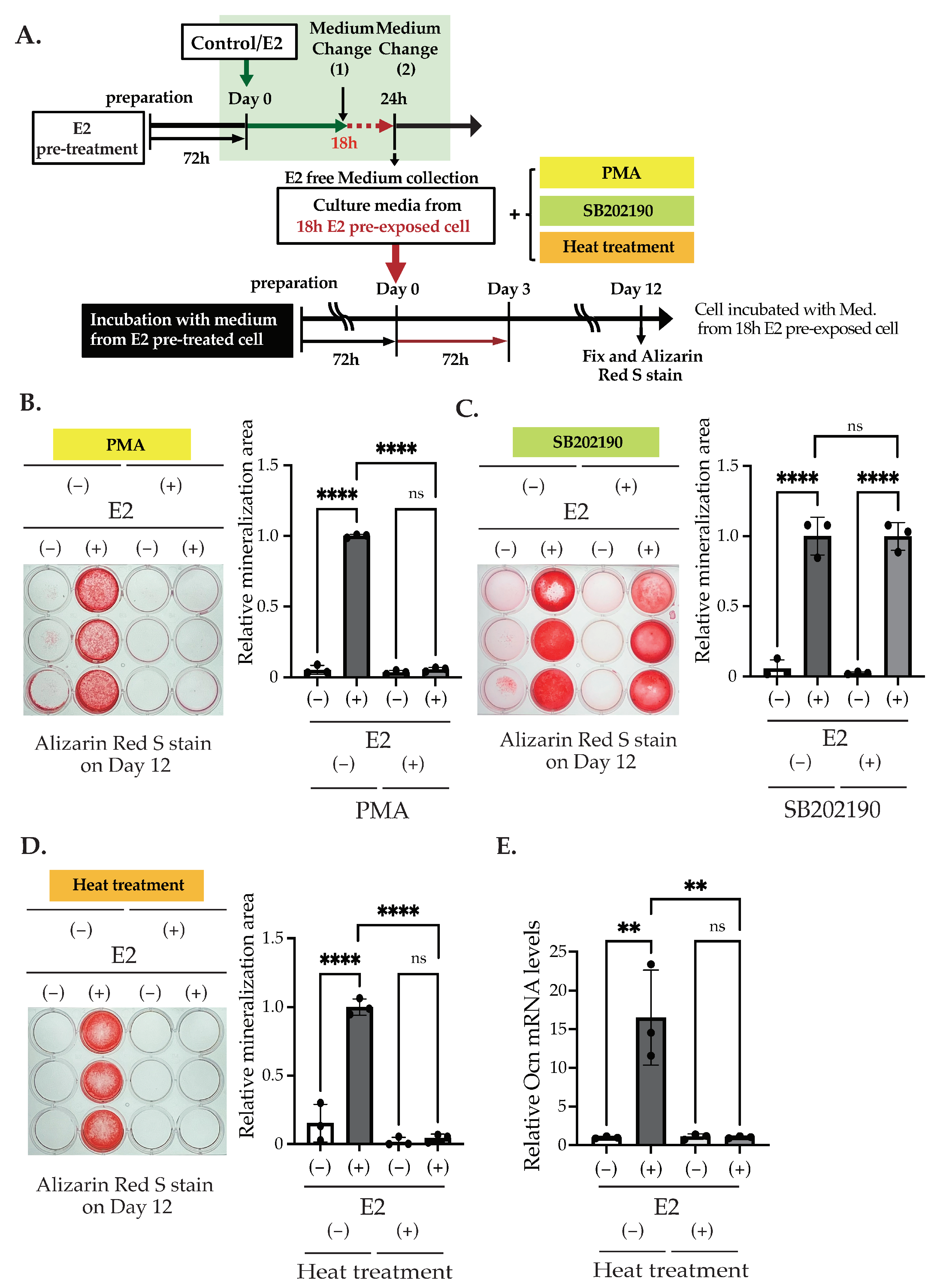
2.3. Inhibition of the p38 MAPK Pathway and the Activation of the PKC Pathway, Excluding the PI3K Pathway, Suppressed E2-Stimulated Matrix Mineralization
2.4. E2 Stimulation Induces p38 MAPK Activation, Which May Inhibit PKC Pathway in an E2- Independent Manner
3. Discussion
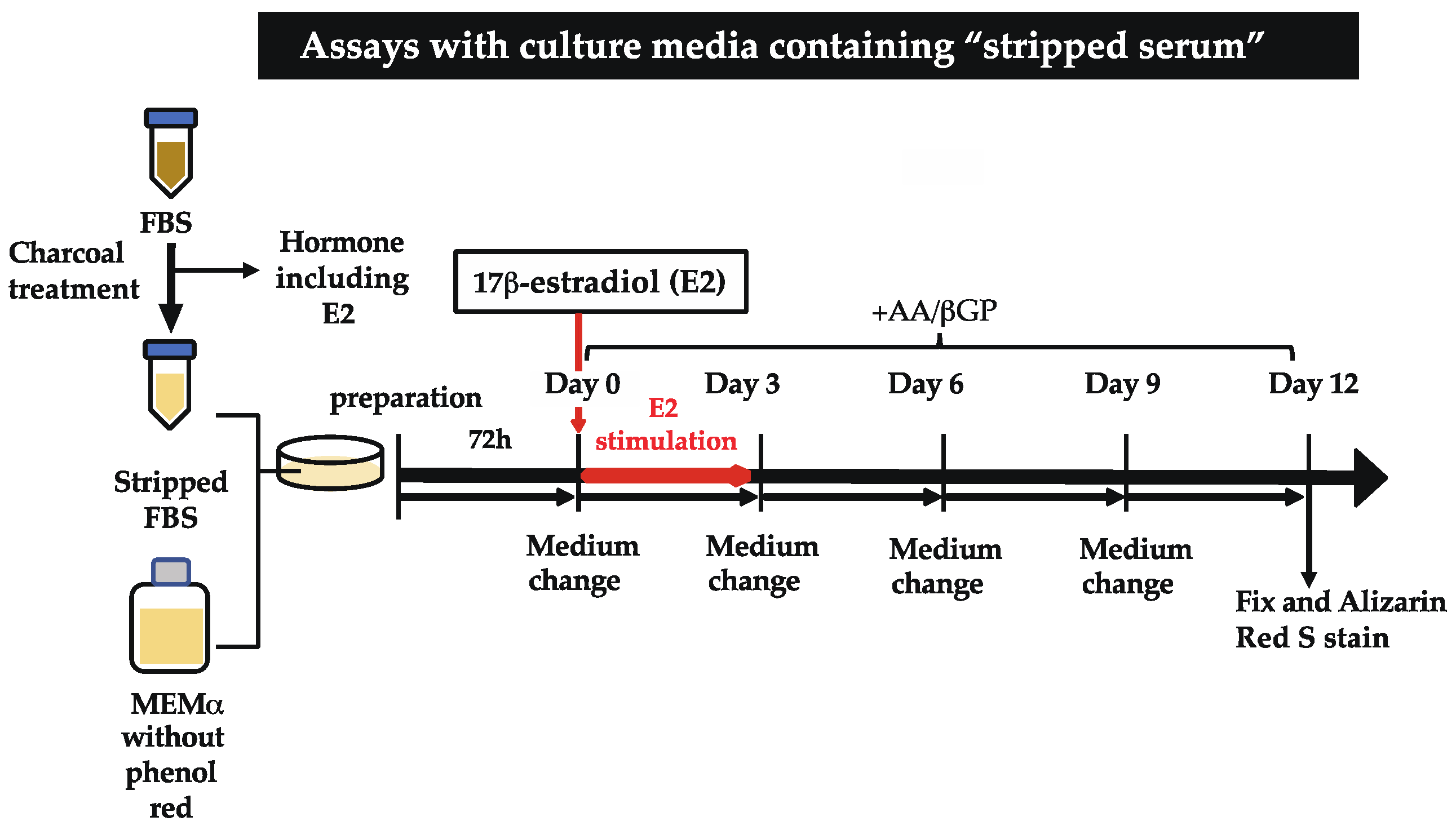
4. Materials and Methods
4.1. Reagents
4.2. Generation of the Stripped Serum
4.3. Cell Line and Culture
4.4. Alizarin Red S Staining
4.5. Quantitative RT-PCR (qRT-PCR)
4.6. Alkaline Phosphatase (ALP) Assay
4.7. Knockdown of ERs by RNA Interference
4.8. Western Blot Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tella, S.H.; Gallagher, J.C. Prevention and treatment of postmenopausal osteoporosis. J. Steroid Biochem. Mol. Biol. 2014, 142, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Hofbauer, L.C. Osteoporosis treatment: Recent developments and ongoing challenge. Lancet Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, B.; Pressman, A.; Sklarin, P.; Bauer, D.C. Associations between Low Levels of Serum Estradiol, Bone Density, and Fractures among Elderly Women: The Study of Osteoporotic Fractures. J. Clin. Endocrinol. Metab. 1998, 83, 2239–2243. [Google Scholar] [CrossRef] [PubMed]
- Ukon, Y.; Makino, T.; Kodama, J.; Tsukazaki, H.; Tateiwa, D.; Yoshikawa, H.; Kaito, T. Molecular-Based Treatment Strategies for Osteoporosis: A Literature Review. Int. J. Mol. Med. 2019, 20, 2557. [Google Scholar] [CrossRef] [PubMed]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 2012, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Chen, L.R.; Chen, K.H. Osteoporosis Due to Hormone Imbalance: An Overview of the Effects of Estrogen Deficiency and Glucocorticoid Overuse on Bone Turnover. Int. J. Mol. Sci. 2022, 23, 1376. [Google Scholar] [CrossRef] [PubMed]
- Shane, E.; Burr, D.; Abrahamsen, B.; Adler, R.A.; Brown, T.D.; Cheung, A.M.; Cosman, F.; Curtis, J.R.; Dell, R.; Dempster, D.W.; et al. Atypical Subtrochanteric and Diaphyseal Femoral Fractures: Second Report of a Task Force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2014, 29, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Burr, D.; Cauley, J.; Dempster, D.W.; Ebeling, P.R.; Felsenberg, D.; Gagel, R.F.; Gilsanz, V.; Guise, T.; Koka, S.; et al. Bisphosphonate-Associated Osteonecrosis of the Jaw: Report of a Task Force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2007, 22, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef]
- Martin, A.; Xiong, J.; Koromila, T.; Ji, J.S.; Chang, S.; Song, Y.S.; Miller, J.L.; Han, C.Y.; Kostenuik, P.; Krum, S.A.; et al. Estrogens antagonize RUNX2-mediated osteoblast-driven osteoclastogenesis through regulating RANKL membrane association. Bone 2015, 75, 96–104. [Google Scholar] [CrossRef]
- Drake, M.T.; Khosla, S. Hormonal and Systemic Regulation of Sclerostin. Bone 2016, 96, 8–17. [Google Scholar] [CrossRef]
- Hayashi, M.; Nakashima, T.; Yoshimura, N.; Okamoto, K.; Tanaka, S.; Takayanagi, H. Autoregulation of Osteocyte Sema3A Orchestrates Estrogen Action and Counteracts Bone Aging. Cell Metab. 2019, 29, 627–637. [Google Scholar] [CrossRef]
- Okazaki, R.; Inoue, D.; Shibata, M.; Saika, M.; Kido, S.; Ooka, H.; Tomiyama, H.; Sakamoto, Y.; Matsumoto, T. Estrogen promotes early osteoblast differentiation and inhibits adipocyte differentiation in mouse bone marrow stromal cell lines that express estrogen receptor (ER) alpha or beta. Endocrinology 2002, 143, 2349–2356. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Otsuka, F.; Takano-Narazaki, M.; Katsuyama, T.; Nakamura, E.; Tsukamoto, N.; Inagaki, K.; Sada, K.; Makino, H. Estrogen facilitates osteoblast differentiation by upregulating bone morphogenetic protein-4 signaling. Steroids 2013, 78, 513–520. [Google Scholar] [CrossRef]
- Badakhshi, Y.; Shao, W.; Liu, D.; Tian, L.; Pang, J.; Gu, J.; Hu, J.; Jin, T. Estrogen-Wnt signaling cascade regulates expression of hepatic fibroblast growth factor 21. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E292–E304. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, E.; Zhang, H.; Wang, J.; Wu, N.; Chen, X.; Wang, N.; Wen, S.; Nan, G.; Deng, F.; et al. Crosstalk between Wnt/β-Catenin and Estrogen Receptor Signaling Synergistically Promotes Osteogenic Differentiation of Mesenchymal Progenitor Cells. PLoS ONE 2013, 8, e82436. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, Y.; Nagata, M.; Kozloff, K.M.; Welch, J.D.; Mizuhashi, K.; Tokavanich, N.; Hallett, S.A.; Link, D.C.; Nagasawa, T.; Ono, W.; et al. A Wnt-mediated transformation of the bone marrow stromal cell identity orchestrates skeletal regeneration. Nat. Commun. 2020, 11, 332. [Google Scholar] [CrossRef] [PubMed]
- Bain, G.; Müller, T.; Wang, X.; Papkoff, J. Activated beta-catenin induces osteoblast differentiation of C3H10T1/2 cells and participates in BMP2 mediated signal transduction. Biochem. Biophys. Res. Commun. 2003, 301, 84–91. [Google Scholar] [CrossRef]
- Matsuzaki, E.; Takahashi-Yanaga, F.; Miwa, Y.; Hirata, M.; Watanabe, Y.; Sato, N.; Morimoto, S.; Hirofuji, T.; Maeda, K.; Sasaguri, T. Differentiation-inducing factor-1 alters canonical Wnt signaling and suppresses alkaline phosphatase expression in osteoblast-like cell lines. J. Bone Miner. Res. 2006, 8, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Grünberg, J.R.; Hammarstedt, A.; Hedjazifar, S.; Smith, U. The Novel Secreted Adipokine WNT1-inducible Signaling Pathway Protein 2 (WISP2) Is a Mesenchymal Cell Activator of Canonical WNT. J. Biol. Chem. 2014, 289, 6899–6907. [Google Scholar] [CrossRef]
- Ronda, A.C.; Buitrago, C.; Boland, R. Role of estrogen receptors, PKC and Src in ERK2 and p38 MAPK signaling triggered by 17β-estradiol in skeletal muscle cells. J. Steroid Biochem. Mol. Biol. 2010, 122, 287–294. [Google Scholar] [CrossRef]
- Nakura, A.; Higuchi, C.; Yoshida, K.; Yoshikawa, H. PKCα suppresses osteoblastic differentiation. Bone 2011, 48, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Endoh, H.; Sasaki, H.; Maruyama, K.; Takeyama, K.; Waga, I.; Shimizu, T.; Kato, S.; Kawashima, H. Rapid activation of MAP kinase by estrogen in the bone cell line. Biochem. Biophys. Res. Commun. 1997, 235, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.R.; Haas, E.; Prossnitz, E.R.; Barton, M. Non-genomic regulation of vascular cell function and growth by estrogen. Mol. Cell. Endocrinol. 2009, 308, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Tatei, K.; Ohshima, N.; Sato, S.; Izumi, T. Regulation of MC3T3-E1 differentiation by actin cytoskeleton through lipid mediators reflecting the cell differentiation stage. Biochem. Biophys. Res. Commun. 2019, 514, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ohshima, N.; Tatei, K.; Taniguchi, T.; Sato, S.; Izumi, T. The role of autonomously secreted PGE2 and its autocrine/paracrine effect on bone matrix mineralization at the different stages of differentiating MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 2020, 524, 929–993. [Google Scholar] [CrossRef]
- de Faria, A.N.; Zancanela, D.C.; Ramos, A.P.; Torqueti, M.R.; Ciancaglini, P. Estrogen and phenol red free medium for osteoblast culture: Study of the mineralization ability. Cytotechnology 2016, 68, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Kim, K. Non-classical genomic estrogen receptor (ER)/specificity protein and ER/activating protein-1 signaling pathways. J. Mol. Endocrinol. 2008, 41, 263–275. [Google Scholar] [CrossRef]
- Björnström, L.; Sjöberg, M. Mechanisms of estrogen receptor signaling: Convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol. 2005, 19, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Ariyani, W.; Miyazaki, W.; Koibuchi, N. A Novel Mechanism of S-equol Action in Neurons and Astrocytes: The Possible Involvement of GPR30/GPER1. Int. J. Mol. Sci. 2019, 20, 5178. [Google Scholar] [CrossRef]
- Chuang, S.C.; Chen, C.H.; Chou, Y.S.; Ho, M.L.; Chang, J.K. G Protein-Coupled Estrogen Receptor Mediates Cell Proliferation through the cAMP/PKA/CREB Pathway in Murine Bone Marrow Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 6490. [Google Scholar] [CrossRef]
- Seval, Y.; Cakmak, H.; Kayisli, U.A.; Arici, A. Estrogen-mediated regulation of p38 mitogen-activated protein kinase in human endometrium. J. Clin. Endocrinol. Metab. 2006, 91, 2349–2357. [Google Scholar] [CrossRef] [PubMed]
- Ewendt, F.; Feger, M.; Föller, M. Myostatin regulates the production of fibroblast growth factor 23 (FGF23) in UMR106 osteoblast-like cells. Pflugers Arch. 2021, 473, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Suttamanatwong, S.; Franceschi, R.T.; Carlson, A.E.; Gopalakrishnan, R. Regulation of matrix Gla protein by parathyroid hormone in MC3T3-E1 osteoblast-like cells involves protein kinase A and extracellular signal-regulated kinase pathways. J. Cell Biochem. 2007, 102, 496–505. [Google Scholar] [CrossRef]
- Zeng, Z.; Jing, D.; Zhang, X.; Duan, Y.; Xue, F. Cyclic mechanical stretch promotes energy metabolism in osteoblast-like cells through an mTOR signaling-associated mechanism. Int. J. Mol. Med. 2015, 36, 947–956. [Google Scholar] [CrossRef]
- Xiao, H.H.; Gao, Q.G.; Zhang, Y.; Wong, K.C.; Dai, Y.; Yao, X.S.; Wong, M.S. Vanillic acid exerts oestrogen-like activities in osteoblast-like UMR 106 cells through MAP kinase (MEK/ERK)-mediated ER signaling pathway. J. Steroid Biochem. Mol. Biol. 2014, 144, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A.; Lobo-Escolar, A.; Mateo, J.; Gil, J.; Ibarz, E.; Gracia, L. Male osteoporosis: A review. World J. Orthop. 2012, 3, 223–234. [Google Scholar] [CrossRef]
- Eiraku, N.; Chiba, N.; Nakamura, T.; Amir, M.S.; Seong, C.H.; Ohnishi, T.; Kusuyama, J.; Noguchi, K.; Matsuguchi, T. BMP9 directly induces rapid GSK3-β phosphorylation in a Wnt-independent manner through class I PI3K-Akt axis in osteoblasts. FASEB J. 2019, 33, 12124–12134. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol (Alias) | Forward Primer | Reverse Primer |
|---|---|---|
| Alp | ACCTTCTCTCCTCCATCCCT | GTGTGTGTGTGTGTCCTGTC |
| Runx2 | GCCCAGGCGTATTTCAGATG | GGTAAAGGTGGCTGGGTAGT |
| Col1a | TGGGCGCGGCTGGTATGAGTTC | ACCCTGCTACGACAACGTGCC |
| Ocn | GCAGAACAGACAAGTCCCAC | ACCTTATTGC CCTCCTGCTT |
| Rpl13a | AGCTTACCTGGGGCGTCTG | ACATTCTTTTCTGCCTGTTTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, H.; Fujiwara, Y.; Ariyani, W.; Amano, I.; Ishii, S.; Ninomiya, A.K.; Sato, S.; Takaoka, A.; Koibuchi, N. 17β-Estradiol (E2) Activates Matrix Mineralization through Genomic/Nongenomic Pathways in MC3T3-E1 Cells. Int. J. Mol. Sci. 2024, 25, 4727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094727
Suzuki H, Fujiwara Y, Ariyani W, Amano I, Ishii S, Ninomiya AK, Sato S, Takaoka A, Koibuchi N. 17β-Estradiol (E2) Activates Matrix Mineralization through Genomic/Nongenomic Pathways in MC3T3-E1 Cells. International Journal of Molecular Sciences. 2024; 25(9):4727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094727
Chicago/Turabian StyleSuzuki, Hiraku, Yuki Fujiwara, Winda Ariyani, Izuki Amano, Sumiyasu Ishii, Ayane Kate Ninomiya, Seiichi Sato, Akinori Takaoka, and Noriyuki Koibuchi. 2024. "17β-Estradiol (E2) Activates Matrix Mineralization through Genomic/Nongenomic Pathways in MC3T3-E1 Cells" International Journal of Molecular Sciences 25, no. 9: 4727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094727