
Antimicrobial Ionic Liquids: Ante-Mortem Mechanisms of Pathogenic EPEC and MRSA Examined by FTIR Spectroscopy

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Determination of Ionic Liquid Minimal Bactericidal Concentrations
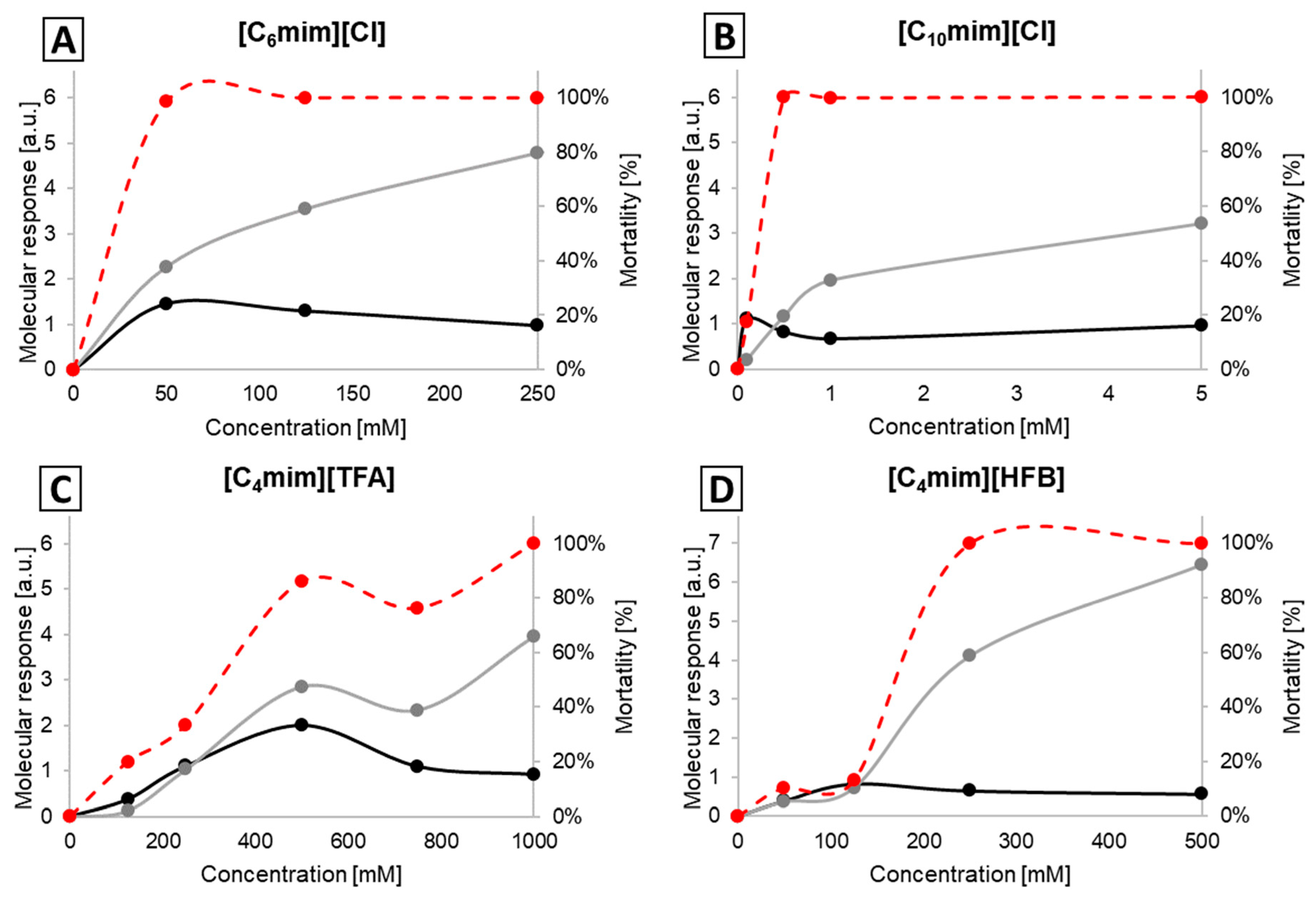
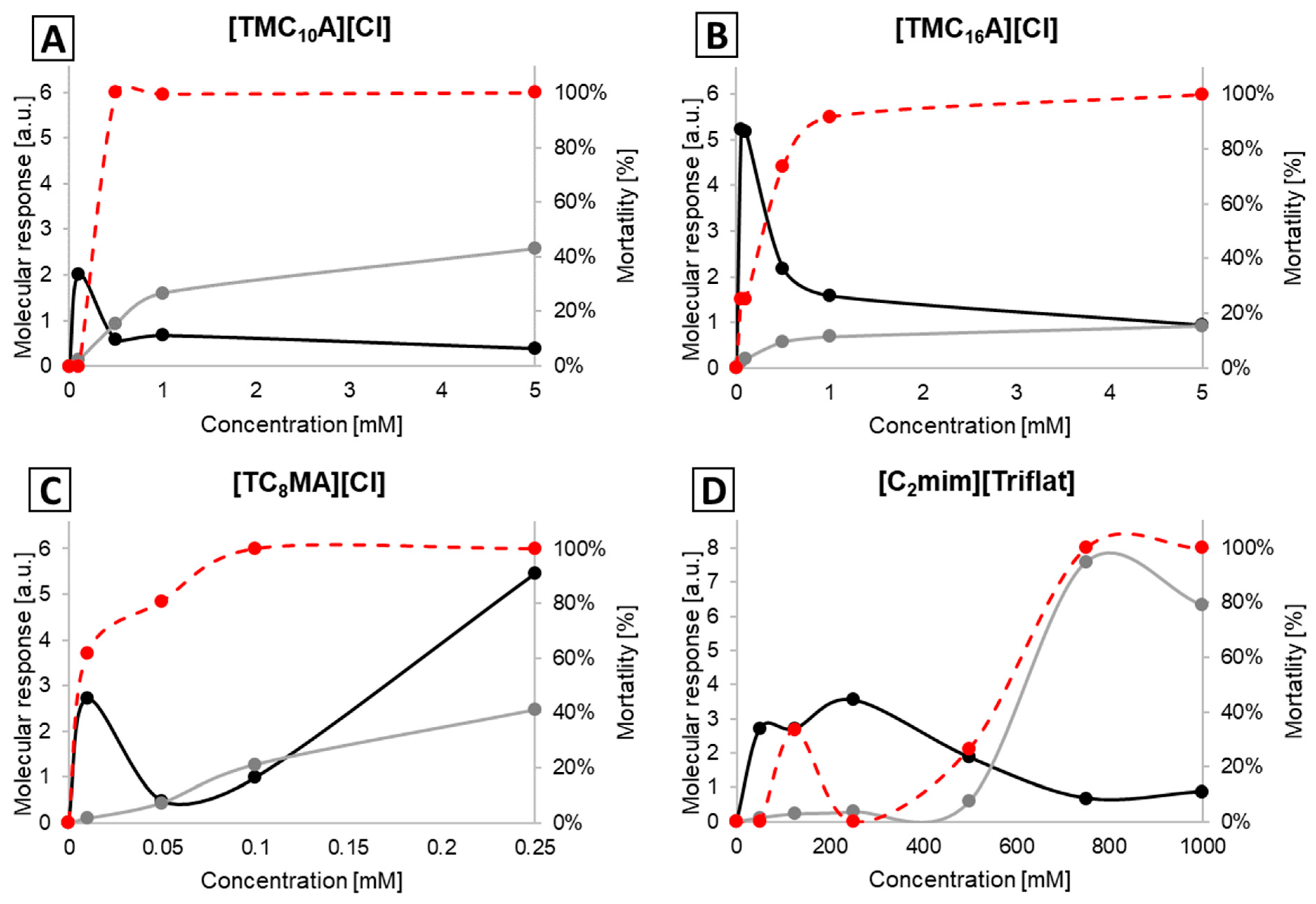
2.2. Molecular Response Analysis for EPEC
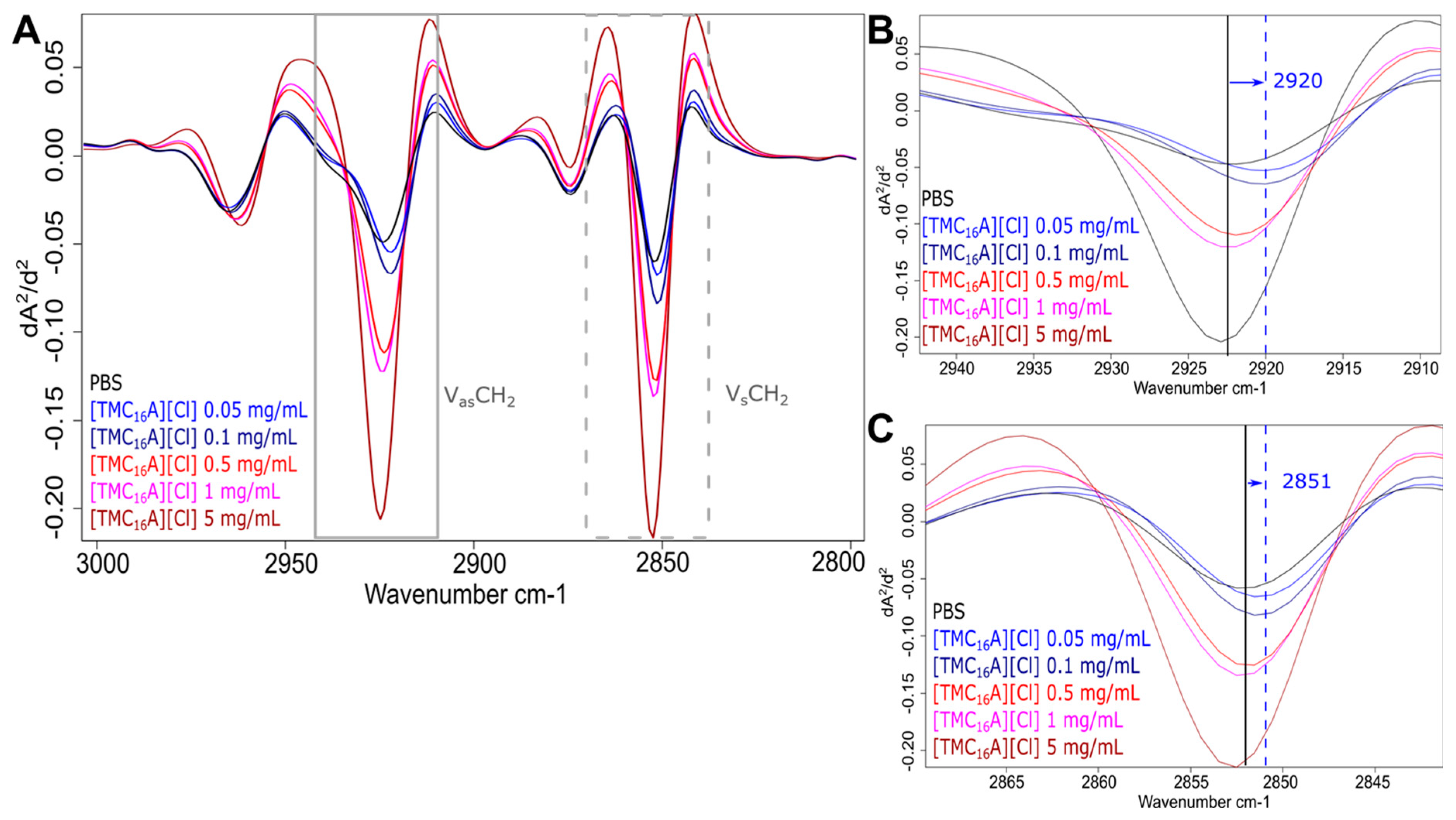
2.3. Molecular Response Analysis for MRSA
3. Materials and Methods
3.1. Ionic Liquids
3.2. Bacterial Strains and Growth Conditions
3.3. FTIR Spectroscopy Procedure
3.4. Spectral Pre-Processing and Molecular Response Analysis
3.5. Biocidal Activity Test
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Egorova, K.S.; Gordeev, E.G.; Ananikov, V.P. Biological Activity of Ionic Liquids and Their Application in Pharmaceutics and Medicine. Chem. Rev. 2017, 117, 7132–7189. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, R.; Branco, L.C.; Prudencio, C.; Noronha, J.P.; Petrovski, Z. Ionic Liquids as Active Pharmaceutical Ingredients. ChemMedChem 2011, 6, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Hough, W.L.; Smiglak, M.; Rodriguez, H.; Swatloski, R.P.; Spear, S.K.; Daly, D.T.; Pernak, J.; Grisel, J.E.; Carliss, R.D.; Soutullo, M.D.; et al. The Third Evolution of Ionic Liquids: Active Pharmaceutical Ingredients. New J. Chem. 2007, 31, 1429–1436. [Google Scholar] [CrossRef]
- Md Moshikur, R.; Chowdhury, M.R.; Moniruzzaman, M.; Goto, M. Biocompatible Ionic Liquids and Their Applications in Pharmaceutics. Green Chem. 2020, 22, 8116–8139. [Google Scholar] [CrossRef]
- Ferraz, R.; Silva, D.; Dias, A.R.; Dias, V.; Santos, M.M.; Pinheiro, L.; Prudêncio, C.; Noronha, J.P.; Petrovski, Ž.; Branco, L.C. Synthesis and Antibacterial Activity of Ionic Liquids and Organic Salts Based on Penicillin g and Amoxicillin Hydrolysate Derivatives against Resistant Bacteria. Pharmaceutics 2020, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhu, Q.; Chen, Z.; Wu, W.; Lu, Y.; Qi, J. Ionic Liquids as a Useful Tool for Tailoring Active Pharmaceutical Ingredients. J. Control. Release 2021, 338, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Pedro, S.N.; Freire, C.S.R.; Silvestre, A.J.D.; Freire, M.G. The Role of Ionic Liquids in the Pharmaceutical Field: An Overview of Relevant Applications. Int. J. Mol. Sci. 2020, 21, 8298. [Google Scholar] [CrossRef]
- Weyhing-Zerrer, N.; Gundolf, T.; Kalb, R.; Oßmer, R.; Rossmanith, P.; Mester, P. Predictability of Ionic Liquid Toxicity from a SAR Study on Different Systematic Levels of Pathogenic Bacteria. Ecotoxicol. Environ. Saf. 2017, 139, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, O.B.; Mutalib, M.I.A.; Leveque, J.M.; El-Harbawi, M. Development of QSAR Model to Predict the Ecotoxicity of Vibrio Fischeri Using COSMO-RS Descriptors. Chemosphere 2017, 170, 242–250. [Google Scholar] [CrossRef]
- Ghanem, O.B.; Papaiconomou, N.; Bdul Mutalib, M.I.; Viboud, S.; El-Harbawi, M.; Uemura, Y.; Gonfa, G.; Azmi Bustam, M.; Leveque, J.M. Thermophysical Properties and Acute Toxicity towards Green Algae and Vibrio Fischeri of Amino Acid-Based Ionic Liquids. J. Mol. Liq. 2015, 212, 352–359. [Google Scholar] [CrossRef]
- Wyrzykowska, E.; Rybińska-Fryca, A.; Sosnowska, A.; Puzyn, T. Virtual Screening in the Design of Ionic Liquids as Environmentally Safe Bactericides. Green Chem. 2019, 21, 1965–1973. [Google Scholar] [CrossRef]
- Mester, P.; Wagner, M.; Rossmanith, P. Antimicrobial Effects of Short Chained Imidazolium-Based Ionic Liquids-Influence of Anion Chaotropicity. Ecotoxicol. Environ. Saf. 2015, 111, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Matzke, M.; Stolte, S.; Thiele, K.; Juffernholz, T.; Arning, J.; Ranke, J.; Welz-Biermann, U.; Jastorff, B. The Influence of Anion Species on the Toxicity of 1-Alkyl-3- Methylimidazolium Ionic Liquids Observed in an (Eco)Toxicological Test Battery. Green Chem. 2007, 9, 1198–1207. [Google Scholar] [CrossRef]
- Stolte, S.; Matzke, M.; Arning, J.; Boschen, A.; Pitner, W.R.; Welz-Biermann, U.; Jastorff, B.; Ranke, J. Effects of Different Head Groups and Functionalised Side Chains on the Aquatic Toxicity of Ionic Liquids. Green Chem. 2007, 9, 1170–1179. [Google Scholar] [CrossRef]
- Kemp, T.J. Ionic Liquids—Pharmaceutical Potential. Sci. Prog. 2012, 95, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Gundolf, T.; Rauch, B.; Kalb, R.; Rossmanith, P.; Mester, P. Influence of Bacterial Lipopolysaccharide Modifications on the Efficacy of Antimicrobial Ionic Liquids. J. Mol. Liq. 2018, 271, 220–227. [Google Scholar] [CrossRef]
- Guo, J.; Qian, Y.; Sun, B.; Sun, Z.; Chen, Z.; Mao, H.; Wang, B.; Yan, F. Antibacterial Amino Acid-Based Poly(Ionic Liquid) Membranes: Effects of Chirality, Chemical Bonding Type, and Application for MRSA Skin Infections. ACS Appl. Bio Mater. 2019, 2, 4418–4426. [Google Scholar] [CrossRef]
- Michalski, J.; Odrzygóźdź, C.; Mester, P.; Narożna, D.; Cłapa, T. Defeat Undefeatable: Ionic Liquids as Novel Antimicrobial Agents. J. Mol. Liq. 2023, 369, 120782. [Google Scholar] [CrossRef]
- Cłapa, T.; Michalski, J.; Syguda, A.; Narożna, D.; van Oostrum, P.; Reimhult, E. Morpholinium-Based Ionic Liquids Show Antimicrobial Activity against Clinical Isolates of Pseudomonas Aeruginosa. Res. Microbiol. 2021, 172, 103817. [Google Scholar] [CrossRef]
- Ferraz, R.; Branco, L.C.; Marrucho, I.M.; Araújo, J.M.M.; Rebelo, L.P.N.; Da Ponte, M.N.; Prudêncio, C.; Noronha, J.P.; Petrovski, E. Development of Novel Ionic Liquids Based on Ampicillin. MedChemComm 2012, 3, 494–497. [Google Scholar] [CrossRef]
- Araújo, J.M.M.; Florindo, C.; Pereiro, A.B.; Vieira, N.S.M.; Matias, A.A.; Duarte, C.M.M.; Rebelo, L.P.N.; Marrucho, I.M. Cholinium-Based Ionic Liquids with Pharmaceutically Active Anions. RSC Adv. 2014, 4, 28126–28132. [Google Scholar] [CrossRef]
- Cvjetko Bubalo, M.; Radosevic, K.; Radojcic Redovnikovic, I.; Halambek, J.; Gaurina Srcek, V. A Brief Overview of the Potential Environmental Hazards of Ionic Liquids. Ecotoxicol. Environ. Saf. 2014, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Heckenbach, M.E.; Romero, F.N.; Green, M.D.; Halden, R.U. Meta-Analysis of Ionic Liquid Literature and Toxicology. Chemosphere 2016, 150, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, S.; Ahmadi, S.; Zohrabi, P. QSAR Modeling of Toxicities of Ionic Liquids toward Staphylococcus aureus Using SMILES and Graph Invariants. Struct. Chem. 2020, 31, 2257–2270. [Google Scholar] [CrossRef]
- Grzonkowska, M.; Sosnowska, A.; Barycki, M.; Rybinska, A.; Puzyn, T. How the Structure of Ionic Liquid Affects Its Toxicity to Vibrio Fischeri? Chemosphere 2016, 159, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Ranke, J.; Molter, K.; Stock, F.; Bottin-Weber, U.; Poczobutt, J.; Hoffmann, J.; Ondruschka, B.; Filser, J.; Jastorff, B. Biological Effects of Imidazolium Ionic Liquids with Varying Chain Lengths in Acute Vibrio Fischeri and WST-1 Cell Viability Assays. Ecotoxicol. Environ. Saf. 2004, 58, 396–404. [Google Scholar] [CrossRef]
- Weyhing-Zerrer, N.; Kalb, R.; Oßmer, R.; Rossmanith, P.; Mester, P. Evidence of a Reverse Side-Chain Effect of Tris(Pentafluoroethyl)Trifluorophosphate [FAP]-Based Ionic Liquids against Pathogenic Bacteria. Ecotoxicol. Environ. Saf. 2018, 148, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Forero-Doria, O.; Araya-Maturana, R.; Barrientos-Retamal, A.; Morales-Quintana, L.; Guzmán, L. N-Alkylimidazolium Salts Functionalized with p-Coumaric and Cinnamic Acid: A Study of Their Antimicrobial and Antibiofilm Effects. Molecules 2019, 24, 3484. [Google Scholar] [CrossRef]
- Naumann, D.; Helm, D.; Labischinski, H. Microbiological Characterizations by FT-IR Spectroscopy. Nature 1991, 351, 81–82. [Google Scholar] [CrossRef]
- Naumann, D. Encyclopedia of Analytical Chemistry; Wiley: Hoboken, NJ, USA, 2000; pp. 102–131. [Google Scholar]
- Vernocchi, P.; Vannini, L.; Gottardi, D.; Del Chierico, F.; Serrazanetti, D.I.; Ndagijimana, M.; Guerzoni, M.E. Integration of Datasets from Different Analytical Techniques to Assess the Impact of Nutrition on Human Metabolome. Front. Cell. Infect. Microbiol. 2012, 2, 156. [Google Scholar] [CrossRef]
- Ami, D.; Natalello, A.; Gatti-Lafranconi, P.; Lotti, M.; Doglia, S.M. Kinetics of Inclusion Body Formation Studied in Intact Cells by FT-IR Spectroscopy. FEBS Lett. 2005, 579, 3433–3436. [Google Scholar] [CrossRef] [PubMed]
- Grunert, T.; Monahan, A.; Lassnig, C.; Vogl, C.; Müller, M.; Ehling-Schulz, M. Deciphering Host Genotype-Specific Impacts on the Metabolic Fingerprint of Listeria Monocytogenes by FTIR Spectroscopy. PLoS ONE 2014, 9, e115959. [Google Scholar] [CrossRef] [PubMed]
- Wenning, M.; Scherer, S. Identification of Microorganisms by FTIR Spectroscopy: Perspectives and Limitations of the Method. Appl. Microbiol. Biotechnol. 2013, 97, 7111–7120. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Ordez, A.; Halisch, J.; Prieto, M. Changes in Fourier Transform Infrared Spectra of Salmonella Enterica Serovars Typhimurium and Enteritidis after Adaptation to Stressful Growth Conditions. Int. J. Food Microbiol. 2010, 142, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Preisner, O.; Lopes, J.A.; Guiomar, R.; MacHado, J.; Menezes, J.C. Fourier Transform Infrared (FT-IR) Spectroscopy in Bacteriology: Towards a Reference Method for Bacteria Discrimination. Anal. Bioanal. Chem. 2007, 387, 1739–1748. [Google Scholar] [CrossRef] [PubMed]
- Wehrli, P.M.; Lindberg, E.; Svensson, O.; Sparen, A.; Josefson, M.; Dunstan, R.H.; Wold, A.E.; Gottfries, J. Exploring Bacterial Phenotypic Diversity Using Factorial Design and FTIR Multivariate Fingerprinting. J. Chemom. 2014, 28, 681–686. [Google Scholar] [CrossRef]
- Winder, C.L.; Goodacre, R. Comparison of Diffuse-Reflectance Absorbance and Attenuated Total Reflectance FT-IR for the Discrimination of Bacteria. Analyst 2004, 129, 1118–1122. [Google Scholar] [CrossRef] [PubMed]
- Corte, L.; Tiecco, M.; Roscini, L.; De Vincenzi, S.; Colabella, C.; Germani, R.; Tascini, C.; Cardinali, G. FTIR Metabolomic Fingerprint Reveals Different Modes of Action Exerted by Structural Variants of N-Alkyltropinium Bromide Surfactants on Escherichia coli and Listeria Innocua Cells. PLoS ONE 2015, 10, e0115275. [Google Scholar] [CrossRef] [PubMed]
- Mester, P.; Jehle, A.K.; Leeb, C.; Kalb, R.; Grunert, T.; Rossmanith, P. FTIR Metabolomic Fingerprint Reveals Different Modes of Action Exerted by Active Pharmaceutical Ingredient Based Ionic Liquids (API-ILs) on: Salmonella Typhimurium. RSC Adv. 2016, 6, 32220–32227. [Google Scholar] [CrossRef]
- Mester, P.; Robben, C.; Witte, A.K.; Kalb, R.; Ehling-Schulz, M.; Rossmanith, P.; Grunert, T. FTIR Spectroscopy Suggests a Revised Mode of Action for the Cationic Side-Chain Effect of Ionic Liquids. ACS Comb. Sci. 2019, 21, 90–97. [Google Scholar] [CrossRef]
- Bromberger, B.; Sommer, J.; Robben, C.; Trautner, C.; Kalb, R.; Rossmanith, P.; Mester, P.J. Evaluation of the Antimicrobial Activity of Pyrithione-Based Ionic Liquids. Sep. Purif. Technol. 2020, 251, 117309. [Google Scholar] [CrossRef]
- Naumann, D.; Labischinski, H.; Giesbrecht, P. The Characterization of Microorganisms by Fourier-Transform Infrared Spectroscopy (FT-IR). In Modem Techniques for Rapid Microbiological Analysis; Nelson, W.H., Ed.; VCH Publishers: New York, NY, USA, 1991; pp. 43–96. [Google Scholar]
- Helm, D.; Labischinski, H.; Schallehn, G.; Naumann, D. Classification and Identifictaion of Bacteria by Fourier-Transform Infrared Spectroscopy. J. Gen. Microbiol. 1991, 137, 69–79. [Google Scholar]
- Socrates, G. Infrared and Raman Characteristic Group Frequencies: Tables and Charts, 3rd ed.; Wiley: Hoboken, NJ, USA, 2004; ISBN 978-0-470-09307-8. [Google Scholar]
- Miller, L.M.; Bourassa, M.W.; Smith, R.J. FTIR Spectroscopic Imaging of Protein Aggregation in Living Cells. Biochim. Biophys. Acta 2013, 1828, 2339–2346. [Google Scholar] [CrossRef]
- Ibsen, K.N.; Ma, H.; Banerjee, A.; Tanner, E.E.L.; Nangia, S.; Mitragotri, S. Mechanism of Antibacterial Activity of Choline-Based Ionic Liquids (CAGE). ACS Biomater. Sci. Eng. 2018, 4, 2370–2379. [Google Scholar] [CrossRef]
- Kóta, Z.; Debreczeny, M.; Szalontai, B. Separable Contributions of Ordered and Disordered Lipid Fatty Acyl Chain Segments to VCH2 Bands in Model and Biological Membranes: A Fourier Transform Infrared Spectroscopic Study. Biospectroscopy 1999, 5, 169–178. [Google Scholar] [CrossRef]
- Scherber, C.M.; Schottel, J.L.; Aksan, A. Membrane Phase Behavior of Escherichia coli during Desiccation, Rehydration, and Growth Recovery. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 2427–2435. [Google Scholar] [CrossRef]
- Beney, L.; Mille, Y.; Gervais, P. Death of Escherichia coli during Rapid and Severe Dehydration Is Related to Lipid Phase Transition. Appl. Microbiol. Biotechnol. 2004, 65, 457–464. [Google Scholar] [CrossRef]
- Gundolf, T.; Weyhing-Zerrer, N.; Sommer, J.; Kalb, R.; Schoder, D.; Rossmanith, P.; Mester, P. Biological Impact of Ionic Liquids Based on Sustainable Fatty Acid Anions Examined with a Tripartite Test System. ACS Sustain. Chem. Eng. 2019, 7, 15865–15873. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Z.; Mu, Q.; Wu, X.; Zhang, J.; Mao, D.; Luo, Y.; Alvarez, P.J.J. Ionic Liquid Enriches the Antibiotic Resistome, Especially Efflux Pump Genes, before Significantly Affecting Microbial Community Structure. Environ. Sci. Technol. 2020, 54, 4305–4315. [Google Scholar] [CrossRef]
- Scheinpflug, K.; Krylova, O.; Strahl, H. Measurement of Cell Membrane Fluidity by Laurdan GP: Fluorescence Spectroscopy and Microscopy. Methods Mol. Biol. 2017, 1520, 159–174. [Google Scholar]
- Kalb, R.S.; Stepurko, E.N.; Emel’yanenko, V.N.; Verevkin, S.P. Carbonate Based Ionic Liquid Synthesis (CBILS[Registered Sign]): Thermodynamic Analysis. Phys. Chem. Chem. Phys. 2016, 18, 31904–31913. [Google Scholar] [CrossRef]
- Kalb, R.; Wesner, W.; Hermann, R.; Kotschan, M.; Schelch, M.; Staber, W. Verfahren zur Herstellung Ionischer Flüssigkeiten, Ionischer Feststoffe Oder Gemische Derselben. WO2005021484A3, 26 May 2005. [Google Scholar]
- Grunert, T.; Wenning, M.; Barbagelata, M.S.; Fricker, M.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M. Rapid and Reliable Identification of Staphylococcus aureus Capsular Serotypes by Means of Artificial Neural Network-Assisted Fourier Transform Infrared Spectroscopy. J. Clin. Microbiol. 2013, 51, 2261–2266. [Google Scholar] [CrossRef]
- Bruker Manual Opus Spectroscopy Software, Version 6; Bruker: Billerica, MA, USA, 2006; Volume 3, Chapter “Quant”.
- Tidy, R.J.; Lam, V.; Fimognari, N.; Mamo, J.C.; Hackett, M.J. FTIR Studies of the Similarities between Pathology Induced Protein Aggregation in Vivo and Chemically Induced Protein Aggregation Ex Vivo. Vib. Spectrosc. 2017, 91, 68–76. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Staphylococcus aureus (MRSA) | Escherichia coli (EPEC) | |||||
|---|---|---|---|---|---|---|
| MBC [mg/mL] | d-Values for 2935–2915 cm−1 | d-Values for 1690–1620 cm−1 | MBC [mg/mL] | d-Values for 2935–2915 cm−1 | d-Values for 1690–1620 cm−1 | |
| [C2mim][Cl] | >1000 | 0.21 | 0.32 | >1000 | 0.05 | 0.11 |
| [C4mim][Cl] | >1000 | 1.26 | 2.93 | >1000 | 0.08 | 2.14 |
| [C6mim][Cl] | 125 | 1.30 | 3.54 | 250 | 0.11 | 4.97 |
| [C8mim][Cl] | 5 | 1.00 | 2.24 | 50 | 0.15 | 2.94 |
| [C10mim][Cl] | 0.5 | 0.82 | 1.16 | 5 | 0.11 | 2.51 |
| [TMA][Cl] | >1000 | 2.25 | 0.15 | 1000 | 1.83 | 3.32 |
| [TMC4A][Cl] | >1000 | 4.35 | 0.39 | 750 | 0.06 | 3.87 |
| [TMC8A][Cl] | 25 | 1.08 | 1.74 | 50 | 0.09 | 1.55 |
| [TMC10A][Cl] | 5 | 0.40 | 2.57 | 5 | 0.29 | 1.65 |
| [TMC12A][Cl] | 5 | 0.55 | 2.68 | 5 | 0.06 | 1.25 |
| [TMC16A][Cl] | 5 | 0.94 | 0.92 | 1 | 0.44 | 0.73 |
| [TC8MA][Cl] | 0.1 | 0.98 | 1.27 | 0.1 | 1.17 | 1.76 |
| [DC8DMA][Cl] | 1 | 0.97 | 1.23 | 1.25 | 0.39 | 0.65 |
| [C4mim][BF4] | 125 | 1.76 | 2.54 | 125 | 0.08 | 2.32 |
| [C4mim][HFB] | 250 | 0.65 | 4.11 | 250 | 0.05 | 2.92 |
| [C4mim][PF6] | 750 | 0.74 | 2.98 | 500 | 0.14 | 1.15 |
| [C4mim][TFA] | 1000 | 0.92 | 3.95 | 500 | 0.13 | 2.32 |
| [C2mim][Triflat] | 750 | 0.69 | 7.58 | 750 | 0.19 | 9.02 |
| Frequency (cm−1) | Assignment | References |
|---|---|---|
| 3000–2800 | Fatty acid region; dominated by stretching vibrations of functional groups usually present in the fatty acid components of the bacterial membrane | [43] |
| ~2923 | C-H str (asym) of CH2 functional groups in fatty acids | [30,44] |
| ~2852 | C-H str (sym) of CH2 functional groups in fatty acids | [30,44] |
| 1800–1500 | Amide region; dominated by the amide bands of proteins and peptides | [43] |
| ~1638 | Amide I of β-pleated sheet | [30] |
| ~1630–1620 | Changes in β-pleated sheet conformation caused by misfolding and/or protein aggregation | [45,46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikuni-Mester, P.; Robben, C.; Witte, A.K.; Linke, K.; Ehling-Schulz, M.; Rossmanith, P.; Grunert, T. Antimicrobial Ionic Liquids: Ante-Mortem Mechanisms of Pathogenic EPEC and MRSA Examined by FTIR Spectroscopy. Int. J. Mol. Sci. 2024, 25, 4705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094705
Mikuni-Mester P, Robben C, Witte AK, Linke K, Ehling-Schulz M, Rossmanith P, Grunert T. Antimicrobial Ionic Liquids: Ante-Mortem Mechanisms of Pathogenic EPEC and MRSA Examined by FTIR Spectroscopy. International Journal of Molecular Sciences. 2024; 25(9):4705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094705
Chicago/Turabian StyleMikuni-Mester, Patrick, Christian Robben, Anna K. Witte, Kristina Linke, Monika Ehling-Schulz, Peter Rossmanith, and Tom Grunert. 2024. "Antimicrobial Ionic Liquids: Ante-Mortem Mechanisms of Pathogenic EPEC and MRSA Examined by FTIR Spectroscopy" International Journal of Molecular Sciences 25, no. 9: 4705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094705