
Anti-Photodamage Effect of Agaricus blazei Murill Polysaccharide on UVB-Damaged HaCaT Cells

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of ABP on Cell Viability, Inflammatory Cytokine Secretion, and Skin Barrier Function
2.2. Isolation, Purification, and Structural Analysis
2.3. Effects of ABP Components A1 and A2 on Cell Viability and Proliferation
2.4. Effects of A1 and A2 on Mitochondrial Membrane Potential (ΔΨm)
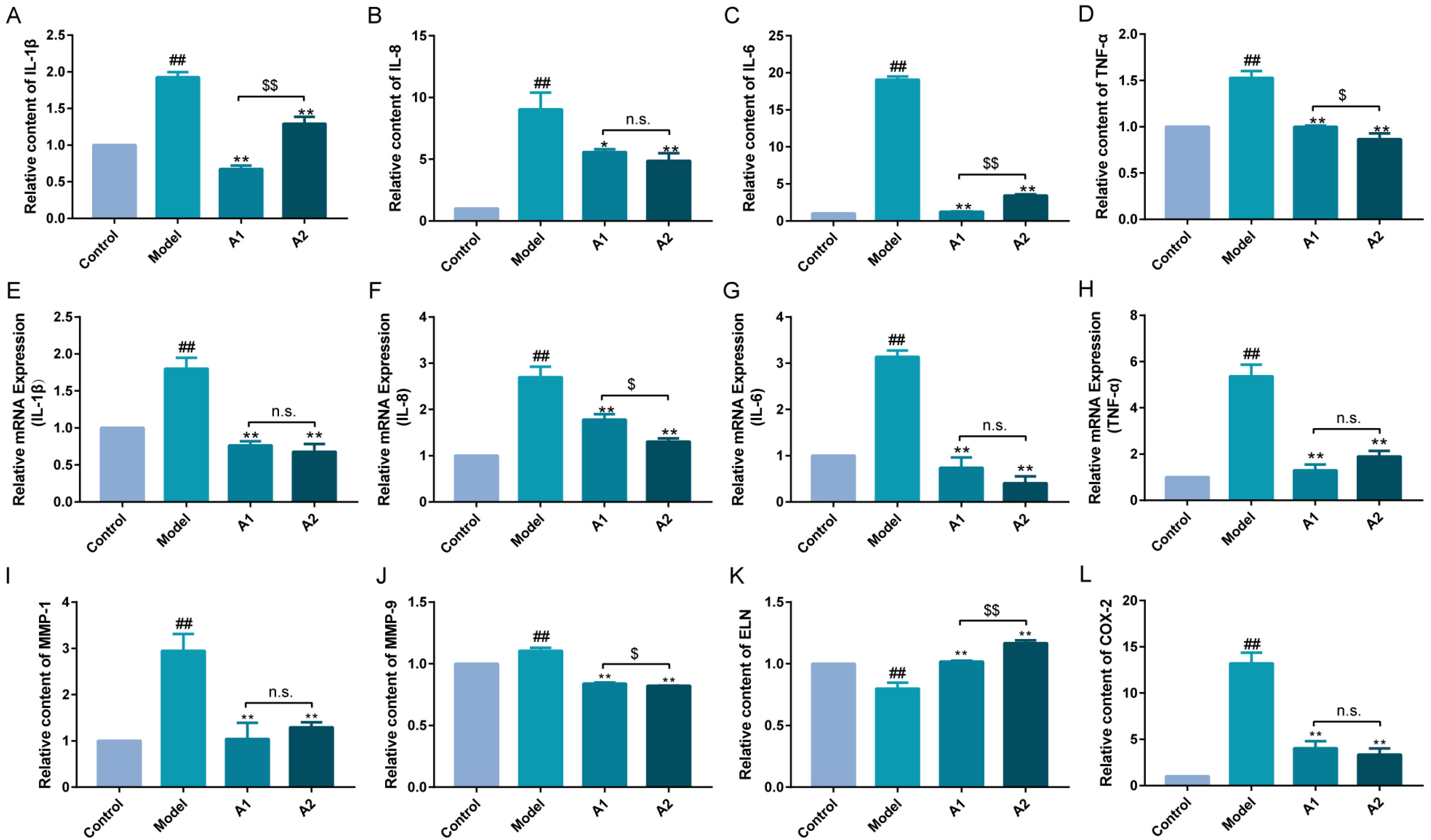
2.5. Effects of A1 and A2 on the Expression Levels of Inflammatory Factors
2.6. Influences of A1 and A2 on MMP-1, MMP-9, ELN, and COX-2
2.7. Expression of JAK-STAT Pathway-Related Genes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction, Separation, and Purification of ABP
4.3. UV–Vis
4.4. FT-IR
4.5. SEM
4.6. AFM
4.7. GPC and Monosaccharide Composition Analysis
4.8. Cell Culture and Toxicity Assay
4.9. The Secretions of Inflammatory Factors and Related Enzymes Were Determined by ELISA
4.10. Determination of Cell Migration Capacity
4.11. Mitochondrial Membrane Potential Measurements
4.12. RT-qPCR
4.13. Immunofluorescence Analysis
4.14. Western Blot
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, T.; Gao, Y.; Zhang, Z.; Li, X.; Zang, P.; Zhao, Y.; He, Z. A study on the anti-skin tumor and anti-UVB damage effects of Gastrodia elata Bl. Products transformed by Armillaria mellea. Food Agric. Immunol. 2022, 33, 736–751. [Google Scholar] [CrossRef]
- Chen, B.; Chen, H.; Qu, H.; Qiao, K.; Xu, M.; Wu, J.; Su, Y.; Shi, Y.; Liu, Z.; Wang, Q. Photoprotective effects of Sargassum thunbergii on ultraviolet B-induced mouse L929 fibroblasts and zebrafish. BMC Complement. Med. Ther. 2022, 22, 144. [Google Scholar] [CrossRef] [PubMed]
- Gunaseelan, S.; Balupillai, A.; Govindasamy, K.; Muthusamy, G.; Ramasamy, K.; Shanmugam, M.; Prasad, N.R. The preventive effect of linalool on acute and chronic UVB-mediated skin carcinogenesis in Swiss albino mice. Photochem. Photobiol. Sci. 2016, 15, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Dare, R.G.; Oliveira, M.M.; Truiti, M.C.T.; Nakamura, C.V.; Ximenes, V.F.; Lautenschlager, S.O.S. Abilities of protocatechuic acid and its alkyl esters, ethyl and heptyl protocatechuates, to counteract UVB-induced oxidative injuries and photoaging in fibroblasts L929 cell line. J. Photochem. Photobiol. B-Biol. 2020, 203, 111771. [Google Scholar] [CrossRef] [PubMed]
- Marionnet, C.; Tricaud, C.; Bernerd, F. Exposure to Non-Extreme Solar UV Daylight: Spectral Characterization, Effects on Skin and Photoprotection. Int. J. Mol. Sci. 2015, 16, 68–90. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, S.; Prasad, N.R.; Karthikeyan, S. Sesamol inhibits UVB-induced ROS generation and subsequent oxidative damage in cultured human skin dermal fibroblasts. Arch. Dermatol. Res. 2010, 302, 733–744. [Google Scholar] [CrossRef]
- Valadares, D.G.; Duarte, M.C.; Ramirez, L.; Chavez-Fumagalli, M.A.; Lage, P.S.; Martins, V.T.; Costa, L.E.; Ribeiro, T.G.; Regis, W.C.B.; Soto, M.; et al. Therapeutic efficacy induced by the oral administration of Agaricus blazei Murill against Leishmania amazonensis. Parasitol. Res. 2012, 111, 1807–1816. [Google Scholar] [CrossRef]
- Ge, Y.; Duan, Y.; Fang, G.; Zhang, Y.; Wang, S. Polysaccharides from fruit calyx of Physalis alkekengi var. francheti: Isolation, purification, structural features and antioxidant activities. Carbohydr. Polym. 2009, 77, 188–193. [Google Scholar] [CrossRef]
- Moradali, M.; Mostafavi, H.; Ghods, S.; Hedjaroude, G. Immunomodulating and anticancer agents in the realm of macromycetes fungi (macrofungi). Int. Immunopharmacol. 2007, 7, 701–724. [Google Scholar] [CrossRef]
- Berven, L.; Karppinen, P.; Hetland, G.; Samuelsen, A.B.C. The Polar High Molecular Weight Fraction of the Agaricus blazei Murill Extract, AndoSan (TM), Reduces the Activity of the Tumor-Associated Protease, Legumain, in RAW 264.7 Cells. J. Med. Food 2015, 18, 429–438. [Google Scholar] [CrossRef]
- Vercosa Junior, D.; Ferraz, V.P.; Duarte, E.R.; Oliveira, N.J.F.; Soto-Blanco, B.; Cassali, G.D.; Melo, M.M. Effects of different extracts of the mushroom Agaricus blazei Murill on the hematologic profile of mice with Ehrlich tumor. Arq. Bras. Med. Vet. Zootec. 2015, 67, 679–688. [Google Scholar] [CrossRef]
- Kasai, H.; He, L.M.; Kawamura, M.; Yang, P.T.; Deng, X.W.; Munkanta, M.; Yamashita, A.; Terunuma, H.; Hirama, M.; Horiuchi, I.; et al. IL-12 Production Induced by Agaricus blazei Fraction H (ABH) Involves Toll-like Receptor (TLR). Evid.-Based Complement Altern. Med. 2004, 1, 259–267. [Google Scholar] [CrossRef]
- Wu, S.; Li, F.; Jia, S.; Ren, H.; Gong, G.; Wang, Y.; Lv, Z.; Liu, Y. Drying effects on the antioxidant properties of polysaccharides obtained from Agaricus blazei Murrill. Carbohydr. Polym. 2014, 103, 414–417. [Google Scholar] [CrossRef]
- Therkelsen, S.P.; Hetland, G.; Lyberg, T.; Lygren, I.; Johnson, E. Cytokine Levels After Consumption of a Medicinal Agaricus blazei Murill-Based Mushroom Extract, AndoSan (TM), in Patients with Crohn’s Disease and Ulcerative Colitis in a Randomized Single-Blinded Placebo-Controlled Study. Scand. J. Immunol. 2016, 84, 323–331. [Google Scholar] [CrossRef]
- Campelo, M.D.S.; Camara Neto, J.F.; Nogueira Lima, A.B.; Das Chagas Neto, F.C.; Da Costa Gonzaga, M.L.; Soares, S.D.A.; Almeida Moreira Leal, L.K.; Nobre Pinho Ribeiro, M.E.; Pontes Silva Ricardo, N.M. Polysaccharides and extracts from Agaricus brasiliensis Murill-A comprehensive review. Int. J. Biol. Macromol. 2021, 183, 1697–1714. [Google Scholar] [CrossRef]
- Davis, J.S.; Ferreira, D.; Paige, E.; Gedye, C.; Boyle, M. Infectious Complications of Biological and Small Molecule Targeted Immunomodulatory Therapies. Clin. Microbiol. Rev. 2020, 33, e00035-19. [Google Scholar] [CrossRef]
- Chen, C.; Fan, N.; Xu, C.; Shao, S.; Shi, G.; Zhou, Y.; Wei, Y.; Wu, L.; Wang, B.; Shi, J.; et al. A synthetic derivative of bioactive constituents from Isatis indigotica ameliorates hypersensitivity and arthritis by inhibiting JAK2-STAT3 pathway in mice. Int. Immunopharmacol. 2023, 124 Pt A, 110884. [Google Scholar] [CrossRef]
- Luo, P.; Ji, Y.; Liu, X.; Zhang, W.; Cheng, R.; Zhang, S.; Qian, X.; Huang, C. Affected inflammation-related signaling pathways in snake envenomation: A recent insight. Toxicon Off. J. Int. Soc. Toxinol. 2023, 234, 107288. [Google Scholar] [CrossRef]
- Luo, Y.; Ali, T.; Liu, Z.; Gao, R.; Li, A.; Yang, C.; Ling, L.; He, L.; Li, S. EPO prevents neuroinflammation and relieves depression via JAK/STAT signaling. Life Sci. 2023, 333, 122102. [Google Scholar] [CrossRef]
- Aghamohammad, S.; Sepehr, A.; Miri, S.T.; Najafi, S.; Pourshafie, M.R.; Rohani, M. Investigation of the anti-inflammatory effects of native potential probiotics as supplementary therapeutic agents in an in-vitro model of inflammation. BMC Complement. Med. Ther. 2023, 23, 335. [Google Scholar] [CrossRef]
- Cheng, W.; An, Q.; Zhang, J.; Shi, X.; Wang, C.; Li, M.; Zhao, D. Protective Effect of Ectoin on UVA/H2O2-Induced Oxidative Damage in Human Skin Fibroblast Cells. Appl. Sci. 2022, 12, 8531. [Google Scholar] [CrossRef]
- Shirley, S.H.; Rundhaug, J.E.; Perez, C.J.; Coletta, L.D.; Kusewitt, D.F. Slug Modulates UV Radiation-Induced Cutaneous Inflammation by Regulating Epidermal Production of Proinflammatory Cytokines. J. Investig. Dermatol. 2017, 137, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; Zou, J.; Rao, Z.; Ji, Y.; Lei, Z.; Peng, L.; Yang, Y.; He, X.; Zeng, N. Polysaccharides from Laminaria japonica: An insight into the current research on structural features and biological properties. Food Funct. 2021, 12, 4254–4283. [Google Scholar] [CrossRef] [PubMed]
- Nie, N.; Bai, C.; Song, S.; Zhang, Y.; Wang, B.; Li, Z. Retraction Note: Bifidobacterium plays a protective role in TNF-α-induced inflammatory response in Caco-2 cell through NF-κB and p38MAPK pathways. Mol. Cell Biochem. 2024, 479, 741. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, Y.; Fu, H.; Yao, F.; Liu, P.; Mo, Q.; Wang, D.; Zhao, D.; Wang, C.; Li, M. Physicochemical and Anti-UVB-Induced Skin Inflammatory Properties of Lacticaseibacillus paracasei Subsp. paracasei SS-01 Strain Exopolysaccharide. Fermentation 2022, 8, 198. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, P.; You, S.; Zhao, D.; An, Q.; Wang, D.; Zhang, J.; Li, M.; Wang, C. Anti-Inflammatory Effects of Opuntia Milpa Alta Polysaccharides Fermented by Lactic Acid Bacteria in Human Keratinocyte HaCaT Cells. Chem. Biodivers. 2022, 19, e202100923. [Google Scholar] [CrossRef]
- Telorac, J.; Prykhozhij, S.V.; Schone, S.; Meierhofer, D.; Sauer, S.; Thomas-Chollier, M.; Meijsing, S.H. Identification and characterization of DNA sequences that prevent glucocorticoid receptor binding to nearby response elements. Nucleic Acids Res. 2016, 44, 6142–6156. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhang, X.; Wu, J. Effect of Gypenosides-containing Serum on Secretion of Inflammatory Factors IL-1β, IL-6 of Photoaging Skin HaCaT Cells. Henan Tradit. Chin. Med. 2013, 33, 48–50. [Google Scholar]
- Pollet, M.; Shaik, S.; Mescher, M.; Frauenstein, K.; Tigges, J.; Braun, S.A.; Sondenheimer, K.; Kaveh, M.; Bruhs, A.; Meller, S.; et al. The AHR represses nucleotide excision repair and apoptosis and contributes to UV-induced skin carcinogenesis (vol 25, pg 1823, 2018). Cell Death Differ. 2020, 27, 403. [Google Scholar] [CrossRef]
- Muthusamy, V.; Hodges, L.D.; Macrides, T.A.; Boyle, G.M.; Piva, T.J. Effect of Novel Marine Nutraceuticals on IL-1α-Mediated TNF-α Release from UVB-Irradiated Human Melanocyte-Derived Cells. Oxidative Med. Cell Longev. 2011, 2011, 728645. [Google Scholar] [CrossRef]
- Wenyu, D.; Linna, F.; Yan, T.; Congfen, H. Study of the protective effects of cosmetic ingredients on the skin barrier, based on the expression of barrier-related genes and cytokines. Mol. Biol. Rep. 2021, 49, 989–995. [Google Scholar]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; Mclean, W.H.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef]
- Hara-Chikuma, M.; Verkman, A.S. Roles of aquaporin-3 in the epidermis. J. Investig. Dermatol. 2008, 128, 2145–2151. [Google Scholar] [CrossRef]
- Verdier-Sevrain, S.; Bonte, F. Skin hydration: A review on its molecular mechanisms. J. Cosmet. Dermatol. 2007, 6, 75–82. [Google Scholar] [CrossRef]
- Zhenzhuo, L.; Rui, J.; Chenxu, J.; Jianzeng, L.; Xiaohao, X.; Liwei, S.; Daqing, Z. Protective effect of oligosaccharides isolated from Panax ginseng C. A. Meyer against UVB-induced skin barrier damage in BALB/c hairless mice and human keratinocytes. J. Ethnopharmacol. 2021, 283, 114677. [Google Scholar]
- Cho, S.M.; Park, J.S.; Kim, K.P.; Cha, D.Y.; Yoo, I.D. Chemical Features and Purification of Immunostimulating Polysaccharides from the Fruit Bodies of Agaricus blazei. Microsurgery 1999, 27, 548–551. [Google Scholar]
- Moon, H.I.; Lee, J.; Kwak, J.H.; Zee, O.P.; Chung, J.H. Isoflavonoid from Viola hondoensis, regulates the expression of matrix metalloproteinase-1 in human skin fibroblasts. Biol. Pharm. Bull. 2005, 28, 925–928. [Google Scholar] [CrossRef]
- Shan, S.; Xiao, T.; Chen, J.; Geng, S.; Li, C.; Xu, X.; Hong, Y.; Ji, C.; Guo, Y.; Wei, H.; et al. Kanglaite attenuates UVB-induced down-regulation of aquaporin-3 in cultured human skin keratinocytes. Int. J. Mol. Med. 2012, 29, 625–629. [Google Scholar] [CrossRef]
- Verkman, A.S. A cautionary note on cosmetics containing ingredients that increase aquaporin-3 expression. Exp. Dermatol. 2008, 17, 871–872. [Google Scholar] [CrossRef]
- Nakakoshi, M.; Morishita, Y.; Usui, K.; Ohtsuki, M.; Ishibashi, K. Identification of a keratinocarcinoma cell line expressing AQP3. Biol. Cell 2006, 98, 95–100. [Google Scholar] [CrossRef]
- Sano, Y.; Masuda, K.; Tamagawa-Mineoka, R.; Matsunaka, H.; Murakami, Y.; Yamashita, R.; Morita, E.; Katoh, N. Thymic stromal lymphopoietin expression is increased in the horny layer of patients with atopic dermatitis. Clin. Exp. Immunol. 2013, 171, 330–337. [Google Scholar] [CrossRef]
- Nylanderlundqvist, E.; Egelrud, T. Formation of active IL-1 beta from pro-IL-1 beta catalyzed by stratum corneum chymotryptic enzyme in vitro. Acta Derm.-Venereol. 1997, 77, 203–206. [Google Scholar] [CrossRef]
- Sun, Q.; Fang, J.; Wang, Z.; Song, Z.; Geng, J.; Wang, D.; Wang, C.; Li, M. Two Laminaria japonica Fermentation Broths Alleviate Oxidative Stress and Inflammatory Response Caused by UVB Damage: Photoprotective and Reparative Effects. Mar. Drugs 2022, 20, 650. [Google Scholar] [CrossRef]
- Chien, R.; Yen, M.; Tseng, Y.; Mau, J. Chemical characteristics and anti-proliferation activities of Ganoderma tsugae polysaccharides. Carbohydr. Polym. 2015, 128, 90–98. [Google Scholar] [CrossRef]
- Shao, P.; Shao, J.; Han, L.; Lv, R.; Sun, P. Separation, preliminary characterization, and moisture-preserving activity of polysaccharides from Ulva fasciata. Int. J. Biol. Macromol. 2015, 72, 924–930. [Google Scholar] [CrossRef]
- Cai, W.; Hu, T.; Bakry, A.M.; Zheng, Z.; Xiao, Y.; Huang, Q. Effect of ultrasound on size, morphology, stability and antioxidant activity of selenium nanoparticles dispersed by a hyperbranched polysaccharide from Lignosus rhinocerotis. Ultrason. Sonochem. 2018, 42, 823–831. [Google Scholar] [CrossRef]
- Asgher, M.; Urooj, Y.; Qamar, S.A.; Khalid, N. Improved exopolysaccharide production from Bacillus licheniformis MS3: Optimization and structural/functional characterization. Int. J. Biol. Macromol. 2020, 151, 984–992. [Google Scholar] [CrossRef]
- Shi, Y.; Xiong, Q.; Wang, X.; Li, X.; Yu, C.; Wu, J.; Yi, J.; Zhao, X.; Xu, Y.; Cui, H. Characterization of a novel purified polysaccharide from the flesh of Cipangopaludina chinensis. Carbohydr. Polym. 2016, 136, 875–883. [Google Scholar] [CrossRef]
- Min, W.; Fang, X.; Wu, T.; Fang, L.; Liu, C.; Wang, J. Characterization and antioxidant activity of an acidic exopolysaccharide from Lactobacillus plantarum JLAU103. J. Biosci. Bioeng. 2019, 127, 758–766. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.; Zhong, C.; Pu, Y.; Yang, Z.; Bao, Y. Structure characteristics and immunomodulatory activities of a polysaccharide RGRP-1b from radix ginseng Rubra. Int. J. Biol. Macromol. 2021, 189, 980–992. [Google Scholar] [CrossRef]
- Zhao, J.; Hu, Y.; Qian, C.; Hussain, M.; Liu, S.; Zhang, A.; He, R.; Sun, P. The Interaction between Mushroom Polysaccharides and Gut Microbiota and Their Effect on Human Health: A Review. Biology 2023, 12, 122. [Google Scholar] [CrossRef]
- Hou, Y.; Ding, X.; Hou, W.; Song, B.; Yan, X. Structure elucidation and antitumor activity of a new polysaccharide from Maerkang Tricholoma matsutake. Int. J. Biol. Sci. 2017, 13, 935–948. [Google Scholar] [CrossRef]
- Li, S.; Shah, N.P. Characterization, antioxidative and bifidogenic effects of polysaccharides from Pleurotus eryngii after heat treatments. Food Chem. 2016, 197, 240–249. [Google Scholar] [CrossRef]
- Kang, Q.; Chen, S.; Li, S.; Wang, B.; Liu, X.; Hao, L.; Lu, J. Comparison on characterization and antioxidant activity of polysaccharides from Ganoderma lucidum by ultrasound and conventional extraction. Int. J. Biol. Macromol. 2019, 124, 1137–1144. [Google Scholar] [CrossRef]
- Vogel, V.; Olari, L.; Jachmann, M.; Reich, S.J.; Haering, M.; Kissmann, A.; Rosenau, F.; Riedel, C.U.; Muench, J.; Spellerberg, B. The bacteriocin Angicin interferes with bacterial membrane integrity through interaction with the mannose phosphotransferase system. Front. Microbiol. 2022, 13, 991145. [Google Scholar] [CrossRef]
- Liao, S.; Liang, C.; Ho, M.; Hsu, T.; Tsai, T.; Hsieh, Y.S.Y.; Tsai, C.; Li, S.; Cheng, Y.; Tsao, S.; et al. Immunization of fucose-containing polysaccharides from Reishi mushroom induces antibodies to tumor-associated Globo H-series epitopes. Proc. Natl. Acad. Sci. USA 2013, 110, 13809–13814. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Khoo, K.H.; Chen, S.T.; Lin, C.C.; Wong, C.H.; Lin, C.H. Studies on the immuno-modulating and antitumor activities of Ganoderma lucidum (Reishi) polysaccharides: Functional and proteomic analyses of a fucose-containing glycoprotein fraction responsible for the activities. Bioorg. Med. Chem. 2002, 10, 1057–1062. [Google Scholar] [CrossRef]
- Farcas, A.D.; Mot, A.C.; Parvu, A.E.; Al Toma, V.; Popa, M.A.; Mihai, M.C.; Sevastre, B.; Roman, I.; Vlase, L.; Parvu, M. In Vivo Pharmacological and Anti-inflammatory Evaluation of Xerophyte Plantago sempervirens Crantz. Oxidative Med. Cell Longev. 2019, 2019, 5049643. [Google Scholar] [CrossRef]
- Bu, H.; He, X.; Zhang, Z.; Yin, Q.; Yu, H.; Li, Y. A TPGS-incorporating nanoemulsion of paclitaxel circumvents drug resistance in breast cancer. Int. J. Pharm. 2014, 471, 206–213. [Google Scholar] [CrossRef]
- Xiao, D.; Yin, S.; Fang, Z.; Xu, D.; Xu, J.; Gu, X.; Wang, G.; Li, J. Reduction-sensitive mixed micelles for selective intracellular drug delivery to tumor cells and reversal of multidrug resistance. Int. J. Pharm. 2018, 550, S01850509325. [Google Scholar]
- Kushairi, N.; Phan, C.W.; Sabaratnam, V.; David, P.; Naidu, M. Lion’s Mane Mushroom, Hericium erinaceus (Bull.: Fr.) Pers. Suppresses H2O2-Induced Oxidative Damage and LPS-Induced Inflammation in HT22 Hippocampal Neurons and BV2 Microglia. Antioxidants 2019, 8, 261. [Google Scholar] [CrossRef]
- Kou, R.; Xia, B.; Wang, Z.; Li, J.; Yang, J.; Gao, Y.; Yin, X.; Gao, J. Triterpenoids and meroterpenoids from the edible Ganoderma resinaceum and their potential anti-inflammatory, antioxidant and anti-apoptosis activities. Bioorganic Chem. 2022, 121, 105689. [Google Scholar] [CrossRef]
- Latha, S.S.; Naveen, S.; Pradeep, C.K.; Sivaraj, C.; Dinesh, M.G.; Anilakumar, K.R. Toxicity Assessment of Wild Mushrooms from the Western Ghats, India: An in Vitro and Sub-Acute in Vivo Study. Front. Pharmacol. 2018, 9, 90. [Google Scholar] [CrossRef]
- Gobi, V.V.; Rajasankar, S.; Ramkumar, M.; Dhanalakshmi, C.; Manivasagam, T.; Thenmozhi, A.J.; Essa, M.M.; Chidambaram, R.; Kalandar, A. Agaricus blazei extract abrogates rotenone-induced dopamine depletion and motor deficits by its anti-oxidative and anti-inflammatory properties in Parkinsonic mice. Nutr. Neurosci. 2018, 21, 657–666. [Google Scholar] [CrossRef]
- Hetland, G.; Tangen, J.; Mahmood, F.; Mirlashari, M.R.; Nissen-Meyer, L.S.H.; Nentwich, I.; Therkelsen, S.P.; Tjonnfjord, G.E.; Johnson, E. Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies. Nutrients 2020, 12, 1339. [Google Scholar] [CrossRef]
- Hetland, G.; Johnson, E.; Lyberg, T.; Kvalheim, G. The Mushroom Agaricus blazei Murill Elicits Medicinal Effects on Tumor, Infection, Allergy, and Inflammation through Its Modulation of Innate Immunity and Amelioration of Th1/Th2 Imbalance and Inflammation. Adv. Pharmacol. Sci. 2011, 2011, 157015. [Google Scholar]
- Frangogiannis, N.G.; Kovacic, J.C. Extracellular Matrix in Ischemic Heart Disease, Part 4/4 JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 2219–2235. [Google Scholar] [CrossRef]
- Balko, J.M.; Schwarz, L.J.; Luo, N.; Estrada, M.V.; Giltnane, J.M.; Davila-Gonzalez, D.; Wang, K.; Sanchez, V.; Dean, P.T.; Combs, S.E.; et al. Triple-negative breast cancers with amplification of JAK2 at the 9p24 locus demonstrate JAK2-specific dependence. Sci. Transl. Med. 2016, 8, 334ra53. [Google Scholar] [CrossRef]
- Qin, X.; Chen, C.; Zhang, Y.; Zhang, L.; Mei, Y.; Long, X.; Tan, R.; Liang, W.; Sun, L. Acitretin modulates HaCaT cells proliferation through STAT1-and STAT3-dependent signaling. Saudi Pharm. J. 2017, 25, 620–624. [Google Scholar] [CrossRef]
- Mittraphab, Y.; Nagata, M.; Matsumoto, M.; Shimizu, K. Antioxidant and Protective Effect of Acetone Extract of Entada phaseoloides Leaves on UVB-Irradiated Human Epidermal Keratinocytes (HaCaT cells) by Inhibiting COX-2, iNOS, and Caspase-3 Activation. Nat. Prod. Commun. 2022, 17, 1934578X221078627. [Google Scholar] [CrossRef]
- Zou, W.; Gonzalez, A.; Jampaiah, D.; Ramanathan, R.; Taha, M.; Walia, S.; Sriram, S.; Bhaskaran, M.; Dominguez-Vera, J.M.; Bansal, V. Skin color-specific and spectrally-selective naked-eye dosimetry of UVA, B and C radiations. Nat. Commun. 2018, 9, 3743. [Google Scholar] [CrossRef]
- Chen, J.; Callac, P.; Parra, L.A.; Karunarathna, S.C.; He, M.Q.; Moinard, M.; De Kesel, A.; Raspe, O.; Wisitrassameewong, K.; Hyde, K.D.; et al. Study in Agaricus subgenus Minores and allied clades reveals a new American subgenus and contrasting phylogenetic patterns in Europe and Greater Mekong Subregion. Persoonia 2017, 38, 170–196. [Google Scholar] [CrossRef]
- Stojkovic, D.; Reis, F.S.; Glamoclija, J.; Ciric, A.; Barros, L.; Van Griensven, L.J.L.D.; Ferreira, I.C.F.R.; Sokovic, M. Cultivated strains of Agaricus bisporus and A. brasiliensis: Chemical characterization and evaluation of antioxidant and antimicrobial properties for the final healthy product—Natural preservatives in yoghurt. Food Funct. 2014, 5, 1602–1612. [Google Scholar] [CrossRef]
- Gonzaga, M.; Ricardo, N.; Heatley, F.; Soares, S.D. Isolation and characterization of polysaccharides from Agaricus blazei Murill. Carbohydr. Polym. 2005, 60, 43–49. [Google Scholar] [CrossRef]
- Grinde, B.; Hedand, G.; Johnson, E. Effects on gene expression and viral load of a medicinal extract from Agaricus blazei in patients with chronic hepatitis C infection. Int. Immunopharmacol. 2006, 6, 1311–1314. [Google Scholar] [CrossRef]
- Wang, Y.; Xiong, X.; Huang, G. Ultrasound-assisted extraction and analysis of maidenhairtree polysaccharides. Ultrason. Sonochem. 2023, 95, 106395. [Google Scholar] [CrossRef]
- Li, Q.; Li, S. Isolation and physico-chemical properties of a new proteoglycan from cultured mycelia of Agaricus blazei. Southest China J. Agric. Sci. 2003, 16, 30–33. [Google Scholar]
- Lin, Y. Some Propertis of Water-soluble Polysaccharides from Agaricus blazei Murill Fruit Body. Food Sci. 2002, 23, 31–33. [Google Scholar]
- Vercosa Junior, D.; Faria De Oliveira, N.J.; Duarte, E.R.; Bastos, G.A.; Maia Soares, A.C.; Cassali, G.D.; Soto-Blanco, B.; Melo, M.M. Serum hepatic biochemistry and electrophoretic protein profile of healthy and Ehrlich tumor-bearing mice treated with extracts of Agaricus blazei Murill. Semin.-Cienc. Agrar. 2016, 37, 763–772. [Google Scholar] [CrossRef]
- Wang, W.; Liu, M.; Zhang, M.; Sun, W.; Zhang, J.; Jia, L. Agaricus blazei Murill polysaccharides alleviate oxidative stress and inflammatory responses against liver and lung injury. Food Biosci. 2022, 47, 101645. [Google Scholar] [CrossRef]
- Wang, P.; Li, X.; Sun, L.; Shen, L. Anti-Inflammatory Activity of Water-Soluble Polysaccharide of Agaricus blazei Murill on Ovariectomized Osteopenic Rats. Evid.-Based Complement Altern. Med. 2013, 2013, 164817. [Google Scholar] [CrossRef]
- Long, M.; de la Vega, M.R.; Wen, Q.; Bharara, M.; Jiang, T.; Zhang, R.; Zhou, S.; Wong, P.K.; Wondrak, G.T.; Zheng, H.; et al. An Essential Role of NRF2 in Diabetic Wound Healing. Diabetes 2016, 65, 780–793. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Saw, C.L.L.; Yang, A.Y.; Huang, M.; Liu, Y.; Lee, J.H.; Khor, T.O.; Su, Z.; Shu, L.; Lu, Y.; Conney, A.H.; et al. Nrf2 null enhances UVB-induced skin inflammation and extracellular matrix damages. Cell Biosci. 2014, 4, 39. [Google Scholar] [CrossRef]
- Rehfeldt, S.C.H.; Laufer, S.; Goettert, M.I. A Highly Selective In Vitro JNK3 Inhibitor, FMU200, Restores Mitochondrial Membrane Potential and Reduces Oxidative Stress and Apoptosis in SH-SY5Y Cells. Int. J. Mol. Sci. 2021, 22, 3701. [Google Scholar] [CrossRef]
- Vieira Gomes, D.C.; Oliveira Barros De Alencar, M.V.; Dos Reis, A.C.; Torres De Lima, R.M.; de Oliveira Santos, J.V.; Oliveira Ferreira Da Mata, A.M.; Soares Dias, A.C.; Da Costa Junior, J.S.; Freire De Medeiros, M.D.G.; Correia Jardim Paz, M.F.; et al. Antioxidant, anti-inflammatory and cytotoxic/antitumoral bioactives from the phylum Basidiomycota and their possible mechanisms of action. Biomed. Pharmacother. 2019, 112, 108643. [Google Scholar] [CrossRef]
- Yang, S.; Li, F.; Lu, S.; Ren, L.; Bian, S.; Liu, M.; Zhao, D.; Wang, S.; Wang, J. Ginseng root extract attenuates inflammation by inhibiting the MAPK/NF-κB signaling pathway and activating autophagy and p62-Nrf2-Keap1 signaling in vitro and in vivo. J. Ethnopharmacol. 2022, 283, 114739. [Google Scholar] [CrossRef]
- Farrukh, M.R.; Nissar, U.; Kaiser, P.J.; Afnan, Q.; Sharma, P.R.; Bhushan, S.; Tasduq, S.A. Glycyrrhizic acid (GA) inhibits reactive oxygen Species mediated photodamage by blocking ER stress and MAPK pathway in UV-B irradiated human skin fibroblasts. J. Photochem. Photobiol. B-Biol. 2015, 148, 351–357. [Google Scholar] [CrossRef]
- Xie, D.; Li, Y.; Wang, G.; Jiang, J.; Sun, L. Ultraviolet light-emitting diode irradiation induces reactive oxygen species production and mitochondrial membrane potential reduction in HL-60 cells. J. Int. Med. Res. 2021, 49, 3000605211016623. [Google Scholar] [CrossRef]
- Ragonese, F.; Monarca, L.; De Luca, A.; Mancinelli, L.; Mariani, M.; Corbucci, C.; Gerli, S.; Iannitti, R.G.; Leonardi, L.; Fioretti, B. Resveratrol depolarizes the membrane potential in human granulosa cells and promotes mitochondrial biogenesis. Fertil. Steril. 2021, 115, 1063–1073. [Google Scholar] [CrossRef]
- Distelmaier, F.; Visch, H.; Smeitink, J.A.M.; Mayatepek, E.; Koopman, W.J.H.; Willems, P.H.G.M. The antioxidant Trolox restores mitochondrial membrane potential and Ca2+-stimulated ATP production in human complex I deficiency (vol 87, 515, 2009). J. Mol. Med. 2021, 99, 1173. [Google Scholar] [CrossRef]
- Karin, M.; Clevers, H. Reparative inflammation takes charge of tissue regeneration. Nature 2016, 529, 307–315. [Google Scholar] [CrossRef]
- Wang, S.; Yang, J.; Ding, C.; Li, J.; You, L.; Dai, M.; Zhao, Y. Glutathione S-Transferase Mu-3 Predicts a Better Prognosis and Inhibits Malignant Behavior and Glycolysis in Pancreatic Cancer. Front. Oncol. 2020, 10, 1539. [Google Scholar] [CrossRef]
- Chung, M.G.; Kim, Y.; Cha, Y.K.; Park, T.H.; Kim, Y. Bitter taste receptors protect against skin aging by inhibiting cellular senescence and enhancing wound healing. Nutr. Res. Pract. 2022, 16, 1–13. [Google Scholar] [CrossRef]
- Gendrisch, F.; Esser, P.R.; Schempp, C.M.; Woelfle, U. Luteolin as a modulator of skin aging and inflammation. Biofactors 2021, 47, 170–180. [Google Scholar] [CrossRef]
- Zong, X.; Zhang, H.; Zhu, L.; Deehan, E.C.; Fu, J.; Wang, Y.; Jin, M. Auricularia auricula polysaccharides attenuate obesity in mice through gut commensal Papillibacter cinnamivorans. J. Adv. Res. 2023, 52, 203–218. [Google Scholar] [CrossRef]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. Isolation of components of Streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Zheng, Y.; He, Z.; Guan, P.; He, X.; Hui, L.; Dai, Y. Study of Schiff base formation between dialdehyde cellulose and proteins, and its application for the deproteinization of crude polysaccharide extracts. Ind. Crop. Prod. 2018, 112, 532–540. [Google Scholar] [CrossRef]
- Wei, H.; Shi, Y.; Yuan, Z.; Huang, Z.; Cai, F.; Zhu, J.; Zhang, W.; Li, J.; Xiong, Q.; Wang, Y.; et al. Isolation, Identification, and Anti-Inflammatory Activity of Polysaccharides of Typha angustifolia. Biomacromolecules 2021, 22, 2451–2459. [Google Scholar] [CrossRef]
- Yan, X.; Yan, Z.; Xiong, Q.; Liu, G.; Zhu, J.; Lu, P. Extraction, Purification, Characterization, and Antiangiogenic Activity of Acidic Polysaccharide from Buddleja officinalis. Evid.-Based Complement Altern. Med. 2020, 2020, 5175138. [Google Scholar] [CrossRef]
- Chen, Z.; Yin, C.; Fan, X.; Ma, K.; Yao, F.; Zhou, R.; Shi, D.; Cheng, W.; Gao, H. Characterization of physicochemical and biological properties of Schizophyllum commune polysaccharide extracted with different methods. Int. J. Biol. Macromol. 2020, 156, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Safaryan, M.J.; Ganjloo, A.; Bimakr, M.; Zarringhalami, S. Optimization of Ultrasound-Assisted Extraction, Preliminary Characterization and In Vitro Antioxidant Activity of Polysaccharides from Green Pea Pods. Foods 2016, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, M.; Yang, Z.; Wang, W.; Lin, H.; Xu, S. Selenoprotein S silencing triggers mouse hepatoma cells apoptosis and necrosis involving in intracellular calcium imbalance and ROS-mPTP-ATP. Biochim. Biophys. Acta-Gen. Subj. 2018, 1862, 2113–2123. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A1 | A2 | |
|---|---|---|
| Peak limits (min) | 21.921–24.953 | 25.237–26.727 |
| Mw | 1.5 × 104 | 6.5 × 104 |
| Mz | 2.4 × 104 | 9.1 × 104 |
| Mw/Mn | 1.282 | 1.297 |
| Mz/Mn | 2.011 | 1.818 |
| Monosaccharide Ratio (%) | A1 | A2 |
|---|---|---|
| Mannose | 3.918 | 0.640 |
| Ribose | 0.976 | 0.191 |
| Rhamnose | 3.077 | 0.716 |
| Glucuronic acid | 1.353 | 0.285 |
| Galacturonic acid | 0.617 | 0.571 |
| Glucose | 61.478 | 78.373 |
| Galactose | 21.460 | 11.108 |
| Xylose | —— | 0.977 |
| Arabinose | 5.832 | 6.925 |
| Fucose | 1.290 | 0.215 |
| Total | 100.000 | 100.000 |
| Peak No. | RT | Standard Sugar | Molecular Weight | Standard Curve * |
|---|---|---|---|---|
| 1 | 15.949 | Mannose | 180.16 | y = 29,549.99x |
| 2 | 20.586 | Ribose | 150.13 | y = 35,195.08x |
| 3 | 21.669 | Rhamnose | 164.16 | y = 24,606.19x |
| 4 | 26.147 | Glucuronic acid | 194.14 | y = 23,994.21x |
| 5 | 30.011 | Galacturonic acid | 194.14 | y = 25,574.43x |
| 6 | 31.584 | N-acetyl-glucosamine | 221.21 | y = 21,259.33x |
| 7 | 34.458 | Glucose | 180.16 | y = 28,273.87x |
| 8 | 36.462 | N-acetyl-galactose | 221.21 | y = 27,966.82x |
| 9 | 39.481 | Galactose | 180.16 | y = 33,401.43x |
| 10 | 41.267 | Xylose | 150.13 | y = 37,008.62x |
| 11 | 42.925 | Arabinose | 150.13 | y = 40,595.71x |
| 12 | 48.109 | Fucose | 164.16 | y = 34,153.37x |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, W.; Di, F.; Li, L.; Pu, C.; Wang, C.; Zhang, J. Anti-Photodamage Effect of Agaricus blazei Murill Polysaccharide on UVB-Damaged HaCaT Cells. Int. J. Mol. Sci. 2024, 25, 4676. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094676
Cheng W, Di F, Li L, Pu C, Wang C, Zhang J. Anti-Photodamage Effect of Agaricus blazei Murill Polysaccharide on UVB-Damaged HaCaT Cells. International Journal of Molecular Sciences. 2024; 25(9):4676. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094676
Chicago/Turabian StyleCheng, Wenjing, Feiqian Di, Luyao Li, Chunhong Pu, Changtao Wang, and Jiachan Zhang. 2024. "Anti-Photodamage Effect of Agaricus blazei Murill Polysaccharide on UVB-Damaged HaCaT Cells" International Journal of Molecular Sciences 25, no. 9: 4676. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094676