
Peptides Targeting the IF1–ATP Synthase Complex Modulate the Permeability Transition Pore in Cancer HeLa Cells
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Peptides IF1-O.1 and IF1-O.2 Are Predicted to Target the IF1–OSCP Subunit Interaction
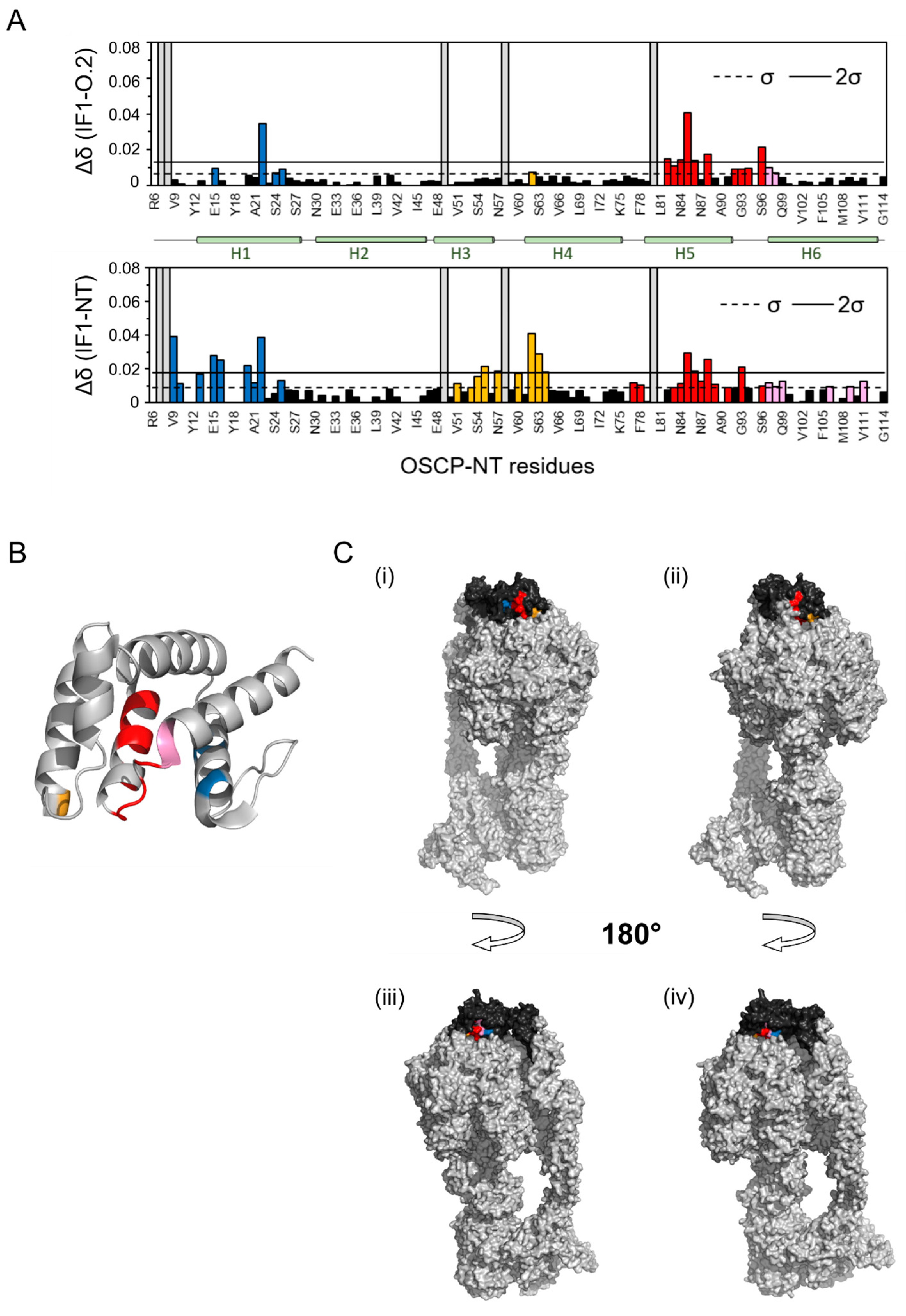
2.2. Peptide IF1-O.1 Displaces IF1 from ATP Synthase and IF1-O.2 Interacts with the IF1 Binding Site on the OSCP Subunit
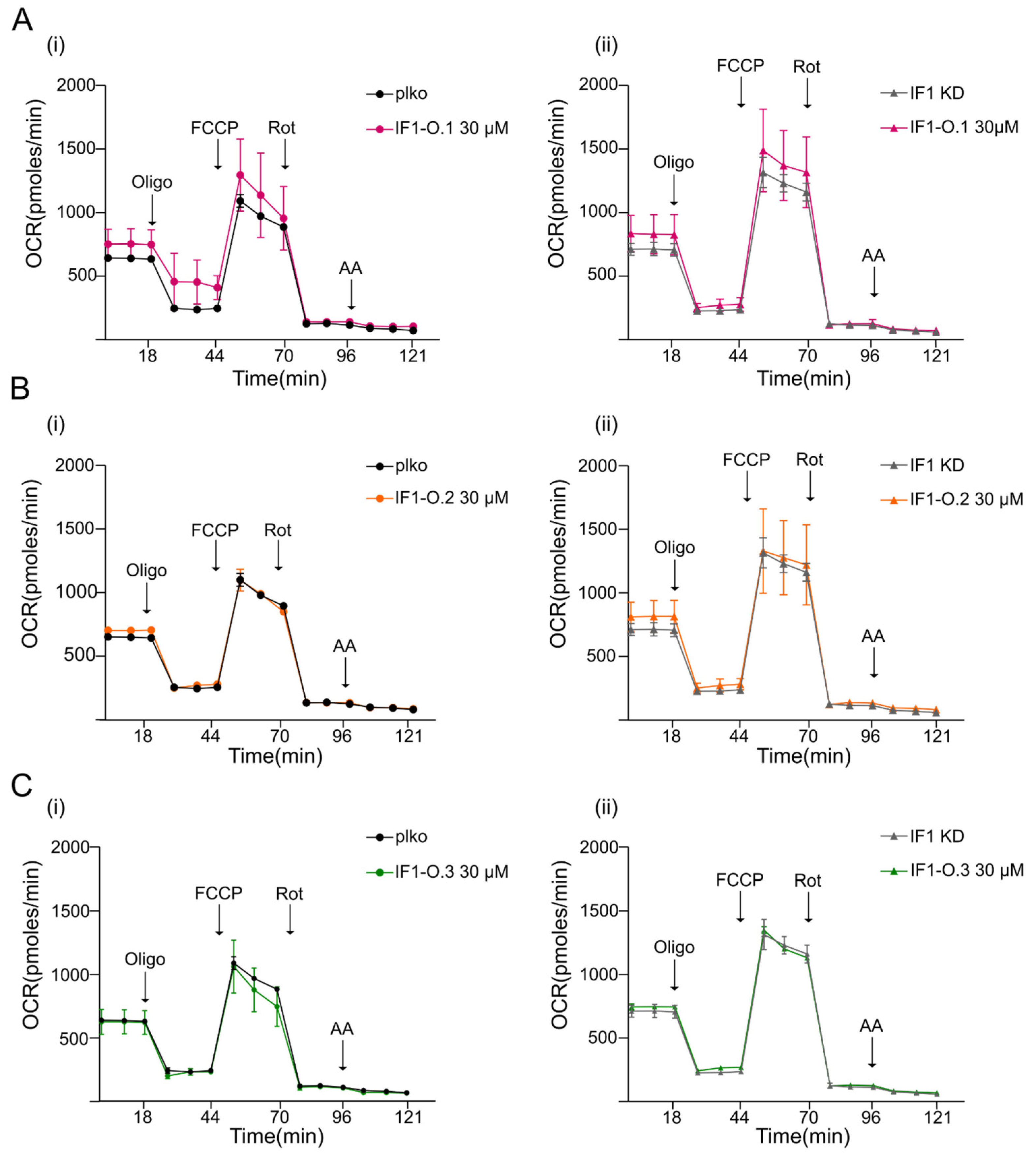
2.3. Peptides IF1-O.1, IF1-O.2 and IF1-O.3 Do Not Affect Mitochondrial Bioenergetics
2.4. Peptides IF1-O.1, IF1-O.2 and IF1-O.3 Modulate the Permeability Transition Pore Opening
3. Discussion
4. Materials and Methods
4.1. Stable Knocking down of IF1 Protein
4.2. Experimental Model
4.3. Peptide Synthesis
4.4. Immunoprecipitation of ATP Synthase
4.5. Nuclear Magnetic Resonance
4.6. Oxygen Consumption Rate
4.7. Calcium Retention Capacity
4.8. Lysates, Gel Electrophoresis, Western Blotting
4.9. Growth Curve
4.10. Quantification and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatto, C.; Grandi, M.; Solaini, G.; Baracca, A.; Giorgio, V. The F1Fo-ATPase Inhibitor Protein IF1 in Pathophysiology. Front. Physiol. 2022, 13, 917203. [Google Scholar] [CrossRef]
- Capuano, F.; Guerrieri, F.; Papa, S. Oxidative Phosphorylation Enzymes in Normal and Neoplastic Cell Growth. J. Bioenerg. Biomembr. 1997, 29, 379–384. [Google Scholar] [CrossRef]
- Bason, J.V.; Montgomery, M.G.; Leslie, A.G.W.; Walker, J.E. Pathway of Binding of the Intrinsically Disordered Mitochondrial Inhibitor Protein to F1-ATPase. Proc. Natl. Acad. Sci. USA 2014, 111, 11305–11310. [Google Scholar] [CrossRef]
- Cabezón, E.; Montgomery, M.G.; Leslie, A.G.W.; Walker, J.E. The Structure of Bovine F1-ATPase in Complex with Its Regulatory Protein IF1. Nat. Struct. Mol. Biol. 2003, 10, 744–750. [Google Scholar] [CrossRef]
- Salewskij, K.; Rieger, B.; Hager, F.; Arroum, T.; Duwe, P.; Villalta, J.; Colgiati, S.; Richter, C.P.; Psathaki, O.E.; Enriquez, J.A.; et al. The Spatio-Temporal Organization of Mitochondrial F1FO ATP Synthase in Cristae Depends on Its Activity Mode. Biochim. Biophys. Acta Bioenerg. 2019, 1861, 148091. [Google Scholar] [CrossRef]
- Green, D.W.; Grover, G.J. The IF1 Inhibitor Protein of the Mitochondrial F1F0-ATPase. Biochim. Biophys. Acta Bioenerg. 2000, 1458, 343–355. [Google Scholar] [CrossRef]
- Gu, J.; Zhang, L.; Zong, S.; Guo, R.; Liu, T.; Yi, J.; Wang, P.; Zhuo, W.; Yang, M. Cryo-EM Structure of the Mammalian ATP Synthase Tetramer Bound with Inhibitory Protein IF1. Science 2019, 364, 1068–1075. [Google Scholar] [CrossRef]
- Galber, C.; Carissimi, S.; Baracca, A.; Giorgio, V. The ATP Synthase Deficiency in Human Diseases. Life 2021, 11, 325. [Google Scholar] [CrossRef]
- Rouslin, W.; Pullman, M.E. Protonic Inhibition o f the Mitochondrial Adenosine 5′-Triphosphatase in Ischemic Cardiac Muscle. Reversible Binding of the ATP Ase Inhibitor Protein to the Mitochondrial ATP Ase during Ischemia. J. Mol. Cell. Cardiol. 1987, 19, 661–668. [Google Scholar] [CrossRef] [PubMed]
- García-Bermúdez, J.; Cuezva, J.M. The ATPase Inhibitory Factor 1 (IF1): A Master Regulator of Energy Metabolism and of Cell Survival. Biochim. Biophys. Acta Bioenerg. 2016, 1857, 1167–1182. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, M.; Imamura, H.; Nakamura, J.; Yoshida, M. Assessing Actual Contribution of IF1, Inhibitor of Mitochondrial FOF1, to ATP Homeostasis, Cell Growth, Mitochondrial Morphology, and Cell Viability. J. Biol. Chem. 2012, 287, 18781–18787. [Google Scholar] [CrossRef] [PubMed]
- Campanella, M.; Casswell, E.; Chong, S.; Farah, Z.; Wieckowski, M.R.; Abramov, A.Y.; Tinker, A.; Duchen, M.R. Regulation of Mitochondrial Structure and Function by the F1FO-ATPase Inhibitor Protein, IF1. Cell Metab. 2008, 8, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Aragó, M.; Formentini, L.; Martínez-Reyes, I.; García-Bermudez, J.; Santacatterina, F.; Sánchez-Cenizo, L.; Willers, I.M.; Aldea, M.; Nájera, L.; Juarránz, Á.; et al. Expression, Regulation and Clinical Relevance of the ATPase Inhibitory Factor 1 in Human Cancers. Oncogenesis 2013, 2, e46. [Google Scholar] [CrossRef]
- Sgarbi, G.; Barbato, S.; Costanzini, A.; Solaini, G.; Baracca, A. The Role of the ATPase Inhibitor Factor 1 (IF1) in Cancer Cells Adaptation to Hypoxia and Anoxia. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 99–109. [Google Scholar] [CrossRef]
- Davies, K.M.; Strauss, M.; Daum, B.; Kief, J.H.; Osiewacz, H.D.; Rycovska, A.; Zickermann, V.; Kuhlbrandt, W. Macromolecular Organization of ATP Synthase and Complex I in Whole Mitochondria. Proc. Natl. Acad. Sci. USA 2011, 108, 14121–14126. [Google Scholar] [CrossRef]
- Blum, T.B.; Hahn, A.; Meier, T.; Davies, K.M.; Kühlbrandt, W. Dimers of Mitochondrial ATP Synthase Induce Membrane Curvature and Self-Assemble into Rows. Proc. Natl. Acad. Sci. USA 2019, 116, 4250–4255. [Google Scholar] [CrossRef] [PubMed]
- Kühlbrandt, W.; Davies, K.M. Rotary ATPases: A New Twist to an Ancient Machine. Trends Biochem. Sci. 2016, 41, 106–116. [Google Scholar] [CrossRef]
- Guo, H.; Rubinstein, J.L. Cryo-EM of ATP Synthases. Curr. Opin. Struct. Biol. 2018, 52, 71–79. [Google Scholar] [CrossRef]
- Walker, J.E. The ATP Synthase: The Understood, the Uncertain and the Unknown. Biochem. Soc. Trans. 2013, 41, 1–16. [Google Scholar] [CrossRef]
- Kahancová, A.; Sklenář, F.; Ježek, P.; Dlasková, A. Overexpression of Native IF1 Downregulates Glucose-Stimulated Insulin Secretion by Pancreatic INS-1E Cells. Sci. Rep. 2020, 10, 1551–1564. [Google Scholar] [CrossRef]
- García-Bermúdez, J.; Sánchez-Aragó, M.; Soldevilla, B.; del Arco, A.; Nuevo-Tapioles, C.; Cuezva, J.M. PKA Phosphorylates the ATPase Inhibitory Factor 1 and Inactivates Its Capacity to Bind and Inhibit the Mitochondrial H+-ATP Synthase. Cell Rep. 2015, 12, 2143–2155. [Google Scholar] [CrossRef]
- Kahancová, A.; Sklenář, F.; Ježek, P.; Dlasková, A. Regulation of Glucose-Stimulated Insulin Secretion by ATPase Inhibitory Factor 1 (IF1). FEBS Lett. 2018, 592, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Caudal, A.; Tang, X.; Chavez, J.D.; McMillen, T.S.; Keller, A.; Villet, O.; Zhao, M.; Liu, Y.; Ritterhoff, J.; et al. Upregulation of Mitochondrial ATPase Inhibitory Factor 1 (ATPIF1) Mediates Increased Glycolysis in Mouse Hearts. J. Clin. Investig. 2022, 132, e155333. [Google Scholar] [CrossRef] [PubMed]
- Galber, C.; Fabbian, S.; Gatto, C.; Grandi, M.; Carissimi, S.; Acosta, M.J.; Sgarbi, G.; Tiso, N.; Argenton, F.; Solaini, G.; et al. The Mitochondrial Inhibitor IF1 Binds to the ATP Synthase OSCP Subunit and Protects Cancer Cells from Apoptosis. Cell Death Dis. 2023, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.; Watt, I.N.; Wright, C.J.; Ding, S.; Fearnley, I.M.; Walker, J.E. The Inhibitor Protein IF1 from Mammalian Mitochondria Inhibits ATP Hydrolysis but Not ATP Synthesis by the ATP Synthase Complex. J. Biol. Chem. 2024, 9258, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, R.; Mori, S.; Ueno, H.; Noji, H. Kinetic Analysis of the Inhibition Mechanism of Bovine Mitochondrial F1-ATPase Inhibitory Protein Using Biochemical Assay. J. Biochem. 2021, 170, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Sgarbi, G.; Righetti, R.; Del Dotto, V.; Grillini, S.; Giorgio, V.; Baracca, A.; Solaini, G. The Pro-Oncogenic Protein IF1 Does Not Contribute to the Warburg Effect and Is Not Regulated by PKA in Cancer Cells. Biochim. Biophys. Acta Mol. Basis Dis. 2024, 1870, 166879. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, R.; Ueno, H.; Okazaki, K.I.; Noji, H. Molecular Mechanism on Forcible Ejection of ATPase Inhibitory Factor 1 from Mitochondrial ATP Synthase. Nat. Commun. 2023, 14, 1682. [Google Scholar] [CrossRef]
- Zulian, A.; Schiavone, M.; Giorgio, V.; Bernardi, P. Forty Years Later: Mitochondria as Therapeutic Targets in Muscle Diseases. Pharmacol. Res. 2016, 113, 563–573. [Google Scholar] [CrossRef]
- Petronilli, V.; Szabò, I.; Zoratti, M. The Inner Mitochondrial Membrane Contains Ion-Conducting Channels Similar to Those Found in Bacteria. FEBS Lett. 1989, 259, 137–143. [Google Scholar] [CrossRef]
- Szabo, I.; Bernardi, P.; Zoratti, M. Modulation of the Mitochondrial Megachannel by Divalent Cations and Protons. J. Biol. Chem. 1992, 267, 2940–2946. [Google Scholar] [CrossRef] [PubMed]
- Galber, C.; Acosta, M.J.; Minervini, G.; Giorgio, V. The Role of Mitochondrial ATP Synthase in Cancer. Biol. Chem. 2020, 401, 1199–1214. [Google Scholar] [CrossRef] [PubMed]
- Hurst, S.; Gonnot, F.; Dia, M.; Crola Da Silva, C.; Gomez, L.; Sheu, S.S. Phosphorylation of Cyclophilin D at Serine 191 Regulates Mitochondrial Permeability Transition Pore Opening and Cell Death after Ischemia-Reperfusion. Cell Death Dis. 2020, 11, 661. [Google Scholar] [CrossRef]
- Giorgio, V.; von Stockum, S.; Antoniel, M.; Fabbro, A.; Fogolari, F.; Forte, M.; Glick, G.D.; Petronilli, V.; Zoratti, M.; Szabó, I.; et al. Dimers of Mitochondrial ATP Synthase form the Permeability Transition Pore. Proc. Natl. Acad. Sci. USA 2013, 110, 5887–5892. [Google Scholar] [CrossRef]
- Giorgio, V.; Fogolari, F.; Lippe, G.; Bernardi, P. OSCP Subunit of Mitochondrial ATP Synthase: Role in Regulation of Enzyme Function and of Its Transition to a Pore. Br. J. Pharmacol. 2019, 176, 4247–4257. [Google Scholar] [CrossRef] [PubMed]
- Gauba, E.; Guo, L.; Du, H. Cyclophilin D Promotes Brain Mitochondrial F1FO ATP Synthase Dysfunction in Aging Mice. J. Alzheimer’s Dis. 2017, 55, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Faccenda, D.; Tan, C.H.; Seraphim, A.; Duchen, M.R.; Campanella, M. IF1 Limits the Apoptotic-Signalling Cascade by Preventing Mitochondrial Remodelling. Cell Death Differ. 2013, 20, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Faccenda, D.; Nakamura, J.; Gorini, G.; Dhoot, G.K.; Piacentini, M.; Yoshida, M.; Campanella, M. Control of Mitochondrial Remodeling by the ATPase Inhibitory Factor 1 Unveils a Pro-Survival Relay via OPA1. Cell Rep. 2017, 18, 1869–1883. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Kong, J.; Xu, W.; Dong, S.; Du, Y.; Yao, C.; Gao, J.; Ke, S.; Wang, S.; Sun, W. Atpase Inhibitory Factor 1 Inhibition Improves the Antitumor of YC-1 against Hepatocellular Carcinoma. Oncol. Lett. 2018, 16, 5230–5236. [Google Scholar] [CrossRef]
- Walker, J.E.; Carroll, J.; He, J. Reply to Bernardi: The Mitochondrial Permeability Transition Pore and the ATP Synthase. Proc. Natl. Acad. Sci. USA 2020, 117, 2745–2746. [Google Scholar] [CrossRef]
- He, J.; Carroll, J.; Ding, S.; Fearnley, I.M.; Walker, J.E. Permeability Transition in Human Mitochondria Persists in the Absence of Peripheral Stalk Subunits of ATP Synthase. Proc. Natl. Acad. Sci. USA 2017, 114, 9086–9091. [Google Scholar] [CrossRef]
- Niedzwiecka, K.; Tisi, R.; Penna, S.; Lichocka, M.; Plochocka, D.; Kucharczyk, R. Two Mutations in Mitochondrial ATP6 Gene of ATP Synthase, Related to Human Cancer, Affect ROS, Calcium Homeostasis and Mitochondrial Permeability Transition in Yeast. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 117–131. [Google Scholar] [CrossRef]
- Bonora, M.; Giorgi, C.; Pinton, P. Molecular Mechanisms and Consequences of Mitochondrial Permeability Transition. Nat. Rev. Mol. Cell Biol. 2022, 23, 266–285. [Google Scholar] [CrossRef] [PubMed]
- Bonora, M.; Bononi, A.; De Marchi, E.; Giorgi, C.; Lebiedzinska, M.; Marchi, S.; Patergnani, S.; Rimessi, A.; Suski, J.M.; Wojtala, A.; et al. Role of the c Subunit of the FO ATP Synthase in Mitochondrial Permeability Transition. Cell Cycle 2013, 12, 674–683. [Google Scholar] [CrossRef]
- Giorgio, V.; Guo, L.; Bassot, C.; Petronilli, V.; Bernardi, P. Calcium and Regulation of the Mitochondrial Permeability Transition. Cell Calcium 2018, 70, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, V.; Burchell, V.; Schiavone, M.; Bassot, C.; Minervini, G.; Petronilli, V.; Argenton, F.; Forte, M.; Tosatto, S.; Lippe, G.; et al. Ca2+ Binding to F-ATP Synthase β Subunit Triggers the Mitochondrial Permeability Transition. EMBO Rep. 2017, 18, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Carraro, M.; Checchetto, V.; Sartori, G.; Kucharczyk, R.; Di Rago, J.P.; Minervini, G.; Franchin, C.; Arrigoni, G.; Giorgio, V.; Petronilli, V.; et al. High-Conductance Channel Formation in Yeast Mitochondria Is Mediated by F-ATP Synthase e and g Subunits. Cell. Physiol. Biochem. 2018, 50, 1840–1855. [Google Scholar] [CrossRef]
- Galber, C.; Minervini, G.; Cannino, G.; Boldrin, F.; Petronilli, V.; Tosatto, S.; Lippe, G.; Giorgio, V. The f Subunit of Human ATP Synthase Is Essential for Normal Mitochondrial Morphology and Permeability Transition. Cell Rep. 2021, 35, 109111. [Google Scholar] [CrossRef] [PubMed]
- Souroujon, M.C.; Mochly-rosen, D. Interactions in Intracellular Signaling. Nat. Biotechnol. 1998, 16, 919–924. [Google Scholar] [CrossRef]
- Cunningham, A.D.; Qvit, N.; Mochly-Rosen, D. Peptides and Peptidomimetics as Regulators of Protein–Protein Interactions. Curr. Opin. Struct. Biol. 2017, 44, 59–66. [Google Scholar] [CrossRef]
- Lai, Y.; Zhang, Y.; Zhou, S.; Xu, J.; Du, Z.; Feng, Z.; Yu, L.; Zhao, Z.; Wang, W.; Tang, Y.; et al. Structure of the Human ATP Synthase. Mol. Cell 2023, 83, 2137–2147. [Google Scholar] [CrossRef] [PubMed]
- Formentini, L.; Sánchez-Aragó, M.; Sánchez-Cenizo, L.; Cuezva, J.M.C. The Mitochondrial ATPase Inhibitory Factor 1 Triggers a ROS-Mediated Retrograde Prosurvival and Proliferative Response. Mol. Cell 2012, 45, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, L.; Hu, X.-G.; Wu, H.-B.; Cui, Y.-H.; Zhang, X.; Wang, Y. ATPase Inhibitory Factor 1 Expression Is an Independent Prognostic Factor in Non-Small Cell Lung Cancer. Am. J. Cancer Res. 2016, 6, 1141–1148. [Google Scholar] [PubMed]
- Wei, S.; Fukuhara, H.; Kawada, C.; Kurabayashi, A.; Furihata, M.; Ogura, S.I.; Inoue, K.; Shuin, T. Silencing of ATPase Inhibitory Factor 1 Inhibits Cell Growth via Cell Cycle Arrest in Bladder Cancer. Pathobiology 2015, 82, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Shan, Q.; Li, P.; Wu, Y.; Xie, J.; Wang, X. ATPase Inhibitory Factor 1 Is a Potential Prognostic Marker for the Migration and Invasion of Glioma. Oncol. Lett. 2015, 10, 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Song, H.; Liang, Y.; Yin, D.; Zhang, H.; Zheng, T.; Wang, J.; Lu, Z.; Song, X.; Pei, T.; et al. Reciprocal Activation between ATPase Inhibitory Factor 1 and NF-ΚB Drives Hepatocellular Carcinoma Angiogenesis and Metastasis. Hepatology 2014, 60, 1659–1673. [Google Scholar] [CrossRef] [PubMed]
- Gledhill, J.R.; Montgomery, M.G.; Leslie, A.G.W.; Walker, J.E. How the Regulatory Protein, IF1, Inhibits F1-ATPase from Bovine Mitochondria. Proc. Natl. Acad. Sci. USA 2007, 104, 15671–15676. [Google Scholar] [CrossRef] [PubMed]
- Carraro, M.; Jones, K.; Sartori, G.; Schiavone, M.; Antonucci, S.; Kucharczyk, R.; di Rago, J.P.; Franchin, C.; Arrigoni, G.; Forte, M.; et al. The Unique Cysteine of F-ATP Synthase OSCP Subunit Participates in Modulation of the Permeability Transition Pore. Cell Rep. 2020, 32, 108095. [Google Scholar] [CrossRef]
- Bonora, M.; Morganti, C.; Morciano, G.; Pedriali, G.; Lebiedzinska-Arciszewska, M.; Aquila, G.; Giorgi, C.; Rizzo, P.; Campo, G.; Ferrari, R.; et al. Mitochondrial Permeability Transition Involves Dissociation of F1FO ATP Synthase Dimers and C-ring Conformation. EMBO Rep. 2017, 18, 1077–1089. [Google Scholar] [CrossRef]
- Dickson, V.K.; Silvester, J.A.; Fearnley, I.M.; Leslie, A.G.W.; Walker, J.E. On the Structure of the Stator of the Mitochondrial ATP Synthase. EMBO J. 2006, 25, 2911–2918. [Google Scholar] [CrossRef]
- Kornfeld, O.S.; Qvit, N.; Haileselassie, B.; Shamloo, M.; Bernardi, P.; Mochly-Rosen, D. Interaction of Mitochondrial Fission Factor with Dynamin Related Protein 1 Governs Physiological Mitochondrial Function In Vivo. Sci. Rep. 2018, 8, 14034. [Google Scholar] [CrossRef] [PubMed]
- Frezza, C.; Cipolat, S.; Scorrano, L. Organelle Isolation: Functional Mitochondria from Mouse Liver, Muscle and Cultured Filroblasts. Nat. Protoc. 2007, 2, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced Software for Biomolecular NMR Spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed]
- Schanda, P.; Brutscher, B. Very Fast Two-Dimensional NMR Spectroscopy for Real-Time Investigation of Dynamic Events in Proteins on the Time Scale of Seconds. J. Am. Chem. Soc. 2005, 127, 8014–8015. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grandi, M.; Fabbian, S.; Solaini, G.; Baracca, A.; Bellanda, M.; Giorgio, V. Peptides Targeting the IF1–ATP Synthase Complex Modulate the Permeability Transition Pore in Cancer HeLa Cells. Int. J. Mol. Sci. 2024, 25, 4655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094655
Grandi M, Fabbian S, Solaini G, Baracca A, Bellanda M, Giorgio V. Peptides Targeting the IF1–ATP Synthase Complex Modulate the Permeability Transition Pore in Cancer HeLa Cells. International Journal of Molecular Sciences. 2024; 25(9):4655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094655
Chicago/Turabian StyleGrandi, Martina, Simone Fabbian, Giancarlo Solaini, Alessandra Baracca, Massimo Bellanda, and Valentina Giorgio. 2024. "Peptides Targeting the IF1–ATP Synthase Complex Modulate the Permeability Transition Pore in Cancer HeLa Cells" International Journal of Molecular Sciences 25, no. 9: 4655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094655