

Molecular Epidemiology of Carbapenem-Resistant Klebsiella aerogenes in Japan

1
Department of Infectious Diseases, Internal Medicine, Tohoku University Graduate School of Medicine, Sendai 980-8575, Japan
2
Department of Bacteriology, BML Inc., Kawagoe 350-1101, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(8), 4494; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084494
Submission received: 5 March 2024
/
Revised: 7 April 2024
/
Accepted: 16 April 2024
/
Published: 19 April 2024
(This article belongs to the Special Issue Applications of Nanomaterials in the Antimicrobial Sector)
Abstract
:Information regarding Klebsiella aerogenes haboring carbapenemase in Japan is limited. A comprehensive nationwide survey was conducted from September 2014 to December 2022, and 67 non-duplicate strains of carbapenem-resistant K. aerogenes were isolated from 57 healthcare facilities in Japan. Through genetic testing and whole-genome sequencing, six strains were found to possess carbapenemases, including imipenemase (IMP)-1, IMP-6, New Delhi metallo-β-lactamase (NDM)-1, and NDM-5. The strain harboring blaNDM-5 was the novel strain ST709, which belongs to the clonal complex of the predominant ST4 in China. The novel integron containing blaIMP-1 featured the oxacillinase-101 gene, which is a previously unreported structure, with an IncN4 plasmid type. However, integrons found in the strains possessing blaIMP-6, which were the most commonly identified, matched those reported domestically in Klebsiella pneumoniae, suggesting the prevalence of identical integrons. Transposons containing blaNDM are similar or identical to the transposon structure of K. aerogenes harboring blaNDM-5 previously reported in Japan, suggesting that the same type of transposon could have been transmitted to K. aerogenes in Japan. This investigation analyzed mobile genetic elements, such as integrons and transposons, to understand the spread of carbapenemases, highlighting the growing challenge of carbapenem-resistant Enterobacterales in Japan and underscoring the critical need for ongoing surveillance to control these pathogens.
1. Introduction
Klebsiella aerogenes is a Gram-negative, rod-shaped, facultative bacteria belonging to the Enterobacterales. K. aerogenes was previously classified under the Enterobacter genus alongside the Enterobacter cloacae complex, which has emerged as a nosocomial pathogen [1]. Whole-genome sequencing (WGS) of multidrug-resistant isolates led to the reclassification of a species into the genus Klebsiella, currently named K. aerogenes [2].
This bacterium, which basically carries the AmpC β-lactamase on its chromosome, was once a main pathogen in nosocomial infections [3]. However, a decreasing trend in its prevalence has been reported since 2010 in Western European countries, such as France. This is due to the dramatic rise in the Escherichia coli pandemic clone O25:H4-ST131, along with Klebsiella pneumoniae and Enterobacter cloacae complex, extended-spectrum β-lactamase (EBSL), and/or carbapenemase-producing strains [4]. The major Enterobacterales that also cause nosocomial infections in China and the United States are E. coli, K. pneumonia, and E. cloacae complex; K. aerogenes is less frequent [5,6].
Gram-negative bacteria, which are the causative agents of nosocomial infections, exhibit resistance to multiple drugs and have a built-in ability to find new ways to acquire resistance [7]. Currently, one of the most concerning drug-resistant organisms is carbapenem-resistant Enterobacterales (CRE), a notable contributing factor of which is the presence of carbapenemase, which propagates through conjugative transfer via plasmids [8]. Globally, carbapenemase-producing K. pneumoniae and E. cloacae complex are the central focus of discussions on CRE [9].
Notwithstanding global trends, in Japan, the scenario is somewhat different. Interestingly, non-carbapenemase-producing K. aerogenes, which is the causative bacterium of CRE infections, continues to be the predominant pathogen in Japan [10]. Epidemiological studies conducted in major Japanese hospitals support this trend [11]. From another point of view, this indicates that carbapenemase-producing K. aerogenes strains are extremely rare or non-existent. Specifically, the National Institute of Infectious Diseases (NIID) reported a total of 1684 cases of CRE infections in 2018, of which 631 (37.5%) were caused by K. aerogenes. However, among the carbapenem-resistant K. aerogenes strains, only two strains (0.32%) were identified as having carbapenemases; specifically, the imipenemase (IMP) type, which is the most common, is considered to be endemic to Japan [10,12]. Although certainly rare in frequency, K. aerogenes has always been associated with poor outcomes in invasive infections, particularly bloodstream infections, and the presence of carbapenemases in this bacterium demands even greater caution [13].
With the attention focused on carbapenemase-producing K. aerogenes, it is notable that K. aerogenes harboring New Delhi metallo-β-lactamase (NDM)-5 was recently reported for the first time in patients from remote areas within the country without any history of overseas travel [14]. This suggests the potential presence of K. aerogenes strains carrying not only the prevalent IMP-type carbapenemase but also an internationally derived NDM type of carbapenemase potentially lurking in the community or community hospitals [15].
In Japan, the definition of CRE within the country was established in September 2014 under the Infectious Diseases Control Law. Since then, many surveys in the country, including our previously conducted CRE nationwide survey, have been conducted primarily based on that definition [16].
In this study, we performed molecular epidemiological analysis of carbapenemase-producing K. aerogenes strains collected from medical institutions nationwide via a testing center, with the aim to elucidate the type and its transmission of carbapenemase genes and clarify the spread of major carbapenemases by analyzing mobile genetic elements, such as integrons and transposons.
2. Results
2.1. Carbapenem-Resistant Klebsiella aerogenes Isolates across Medical Facilities in Japan
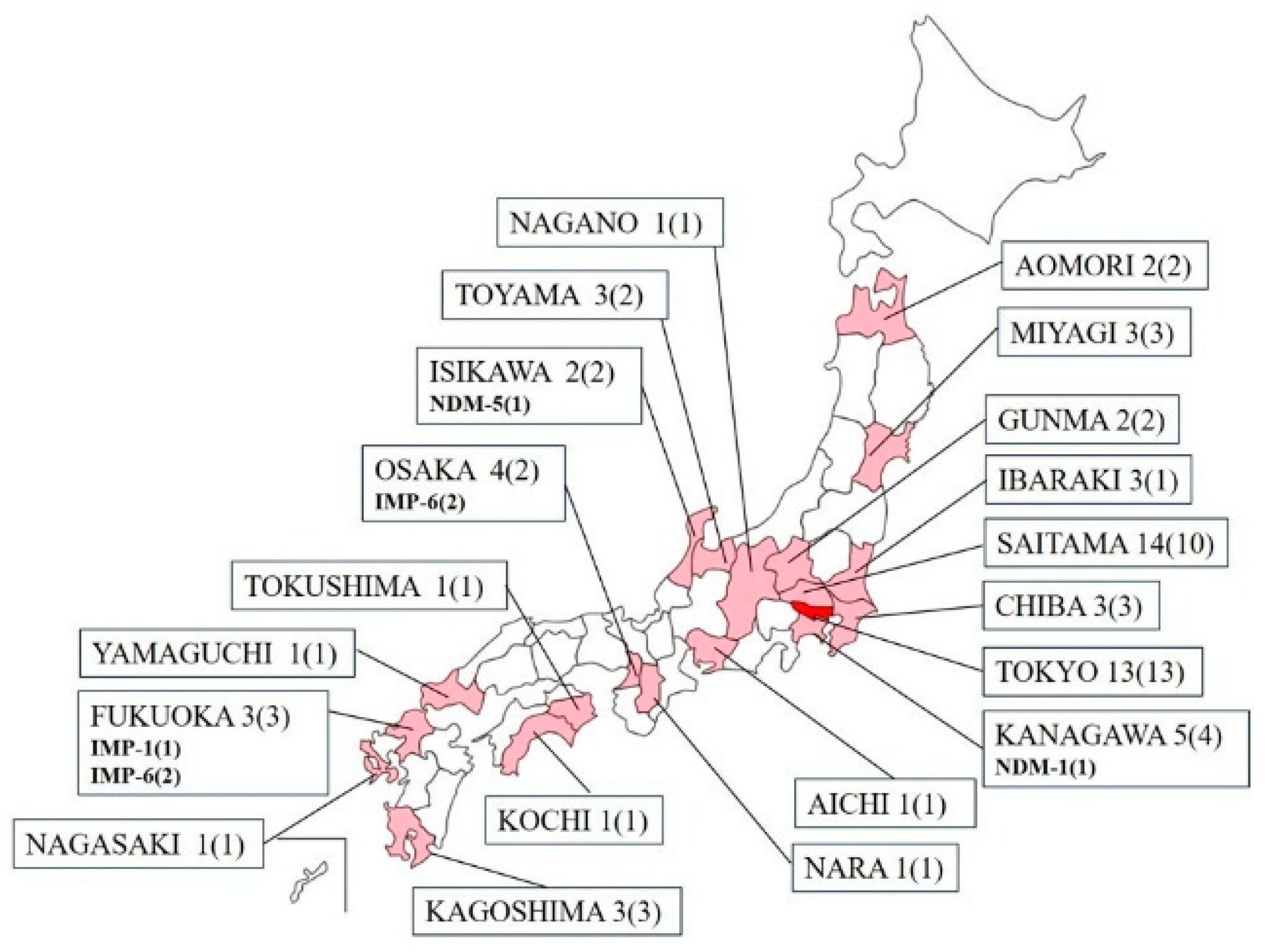
A total of 67 non-duplicate clinical isolates of carbapenem-resistant K. aerogenes from 67 individuals, ranging in age from 0 to 97 years, were collected from 57 medical institutions in 20 prefectures in Japan between September 2014 and December 2022. The Kanto region, including Tokyo, had the largest number of strains, accounting for 59.7% (40/67) of the total, and overall, many of the strains were from eastern Japan (Figure 1). The mean age of the patients from whom each specimen was derived was 75.8 years [95% confidence interval (CI): 71.8–79.9], with a median age of 79 years, including 44 men and 23 women. Among the surveyed medical institutions, the average bed capacity, excluding outpatient clinics, was 198 beds [95% CI: 157–238], with a median bed count of 160. In categorizing the types of healthcare facilities, there were 2 outpatient-only clinics, 41 hospitals that predominantly focused on acute care, 4 dedicated to rehabilitative care, and 10 that catered to long-term or chronic care. Basic information and genetic test results regarding various K. aerogenes strains harboring carbapenemase are demonstrated in Table 1.
2.2. PCR, Whole-Genome Analysis, and MLST
Among the 67 carbapenem-resistant isolates, six tested positive for carbapenemase genes by Polymerase Chain Reaction (PCR), with four testing positive for the IMP gene (blaIMP-1 group) and two for the NDM gene (blaNDM group). Over approximately 8 years, the proportion of carbapenemase-producing K. aerogenes among CRE was 9.0% (6/67). WGS was performed on the six PCR-positive strains, revealing that the carbapenemase-producing genes were IMP-1, IMP-6, NDM-1, and NDM-5 (blaIMP-1, blaIMP-6, blaNDM-1, and blaNDM-5, respectively) using bioinformatic tools. The profiles of the six strains, including those of the other antibiotic resistance genes, are summarized in Table 1.
Referring to the results of drug susceptibility, all strains except TUT0037 were placed on CRE due to resistance to meropenem; TUT0037 was not tested for meropenem susceptibility but was placed on CRE because of drug resistance to imipenem/cilastatin and cefmetazole. Although the strains were observed to harbor the quinolone resistance genes oqxA, dfrA14, and qnrD1, all but TUT0015 showed susceptibility to levofloxacin. TUT0013, TUT0023, and TUT0032 harbored the sul1 gene, but only TUT0023 was resistant to the drug. The results of the drug susceptibility testing for the six strains are listed in the Supplementary Materials (Table S1).
2.3. MLST and Phylogenetic Analysis of Carbapenemase-Producing Klebsiella aerogenes
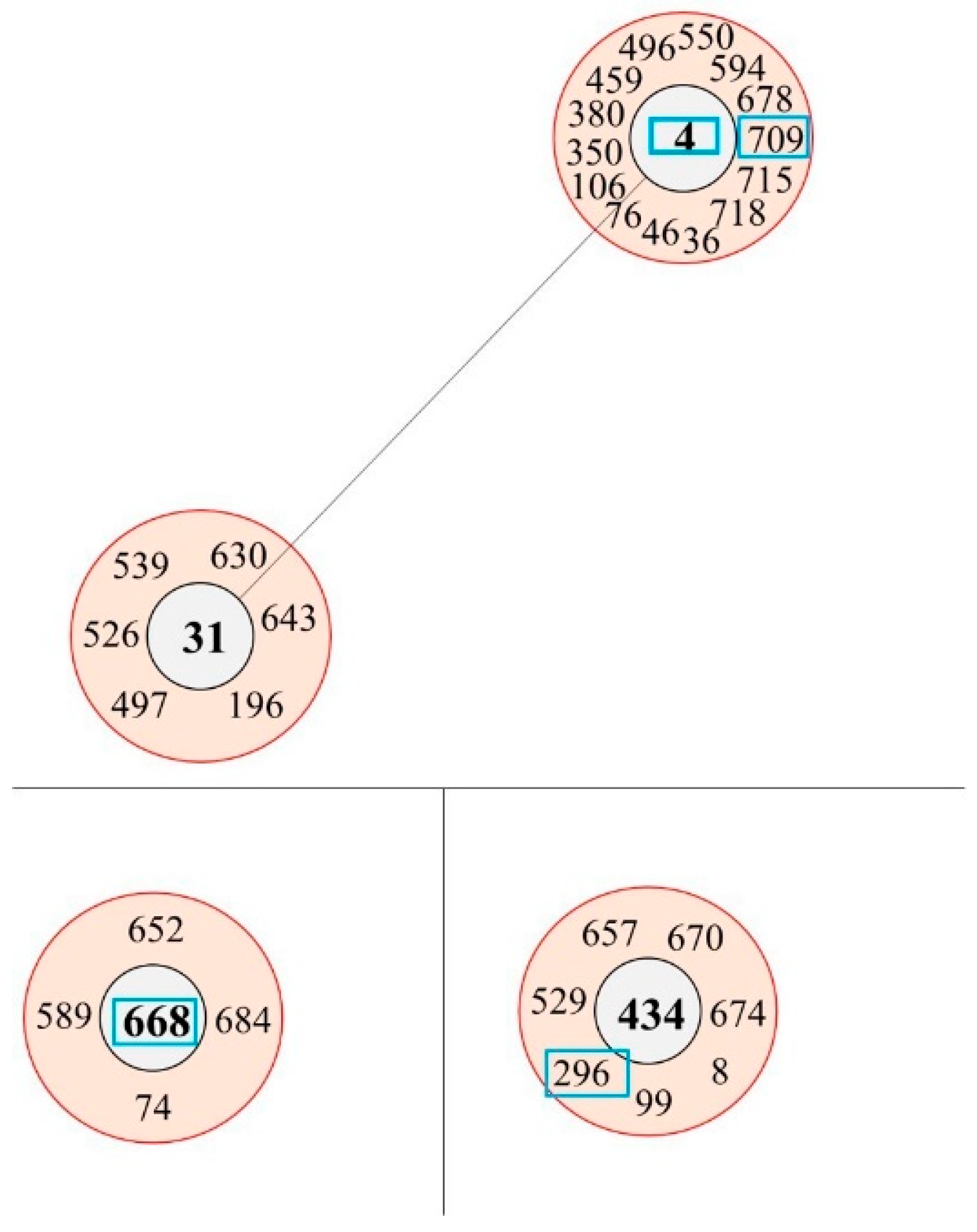
The six K. aerogenes isolates were classified into five STs using multilocus sequence typing (MLST). These belonged to STs 4, 189, 296, 668, and 709. ST709 was a novel detection (Table 1). Two ST668 strains were isolated from different patients in the same hospital, albeit in different years. According to BURST analysis based on allelic profiles, ST4, ST296, ST668, and ST709 were included in three clonal complexes (CCs). ST189 did not constitute a CC. ST4 and ST668 were identified as potential ancestral types (AT); ST296 was included as part of a CC with ST434 as an AT (Figure 2). Overall, there was no uniformity in the STs, and their origins were diverse.
2.4. Plasmid Type (Inc Genes)
The TUT0015 strain was found to have IncN4 on the same contig containing blaIMP-1. Additionally, two strains harboring blaNDM also harbored IncX3. The two strains TUT0023 and TUT0032, which were isolated in Osaka, were discovered to harbor the IncN and IncR genes only through WGS. However, no Inc gene was identified in the contig containing blaIMP-6 for these strains. Similarly, no Inc gene was identified in the contig containing blaIMP-6 for the strain isolated in Fukuoka (TUT0013), even though it had the IncFIA, IncFII, IncH1b, and IncN genes according to WGS.
2.5. Integron Structure Harboring blaIMP
The integron structure was identified in the four strains carrying the blaIMP gene. Of these, three strains had a complete integron structure, while one had a structure known as clusters of attC sites lacking integron integrases (CALIN), which are characterized by the absence of a class 1 integrase gene (intI1) (Figure 3).
Regarding the three strains harboring blaIMP-6, the integron structure of the isolate in Fukuoka in 2016 (TUT0013) consisted of intI1 at the forefront, which was followed downstream by acc(6′)-Ib, blaIMP-6, aadA2, Quaternary ammonium compound resistance protein E delta 1(QacEΔ1), and sul1 with three attC sites (71 bp, 126 bp, and 59 bp). This structure was identified by matching the integron (In1690). The integron of TUT0032 was identical to the integron structure (In1547). Though TUT0023 was missing intI1, the gene cassettes for integrons containing blaIMP-6 were the same for TUT0023 and TUT0032. Unlike the aforementioned three integrons containing blaIMP-6, the integron structure containing blaIMP-1 of the isolate in Fukuoka in 2016 was different. This integron had aph(3′)-XV, blaOXA-101, an efflux small multidrug resistance (SMR) transporter gene, blaIMP-1, and aadA5 with four attC sites (107 bp, 63 bp, 98 bp, and 126 bp) downstream of intl1. Although this integron structure was searched for using Integrall and the NCBI Basic Local Alignment Search Tool (BLAST) for plasmids with a similar cassette structure, no identical structure was identified (Figure 3).
2.6. Transposons Containing blaNDM-1 and blaNDM-5 on IncX3-Type Plasmid
Two strains of K. aerogenes harboring blaNDM were identified in different regions. Strains with blaNDM-5 were isolated in Ishikawa in 2018, and those with blaNDM-1 were isolated in Kanagawa in 2019. Referring to the gene sequence, Tn2, IS3000, and ISAba125 were commonly located upstream of both blaNDM-1 and blaNDM-5, similar to TUT0037 and TUT0049. ISAba125 and IS3000, constituting Tn125 and Tn3000, respectively, were transposases involved in transposon transposition events (Figure 4). However, a paired identical transposase located downstream of the blaNDM was not identified; a composite transposon (CT) could not be confirmed. Genetic abbreviations related to mobile genetic elements are summarized in the Supplemental Materials (Table S2).
3. Discussion
In this study, (1) we conducted a nationwide survey of carbapenem-resistant Klebsiella aerogenes in Japan to determine the frequency of carbapenemase-producing strains, as well as the facilities and regional characteristics from which they were isolated; (2) through whole-genome analysis, we revealed information on prevalent carbapenemase types and STs of K. aerogenes across Japan; and (3) we identified the integrons and transposons encoding carbapenemases and investigated their origins.
3.1. Global and Local Perspectives on Carbapenemase Presence in Klebsiella aerogenes
Information on carbapenemase possession among K. aerogenes strains worldwide is limited. First of all, K. aerogenes has been reported to possess essentially small amounts of carbapenemase. Chromosomal AmpC overproduction and inherent properties have been suggested as possible reasons for the low number of carbapenemase-carrying strains [17]. In a survey of 151 institutions in 35 countries, the rate of carbapenemase possession among multidrug-resistant K. aerogenes strains was reported to be 3.1% [18]. In the two previous reports from China, the positive proportion for carbapenemase ranged from 8.3% to 14.3% among CRE [19,20]. In Japan, the recent NIID annual report shows that carbapenemase-producing K. aerogenes was not reported in some years, and the frequency of its official annual reporting as a cause of infection was estimated to be about 0–0.32% each year [12,21,22,23]. In the current study, the proportion of carbapenemase-producing bacteria among CRE in K. aerogenes was higher than officially reported, accounting for 9%. This may have been due in part to the inclusion of colonization, as well as the bacteria that necessarily caused the infections in this study. Information about the target medical facility is also important and may have contributed to the high carbapenemase proportion. In Japan, 69% of medical institutions with inpatient facilities are small, with less than 200 beds, and these small facilities are often left out of public surveys [24]. The full picture of bacterial resistance in these small healthcare facilities is not known. In this study, the average number of beds in 55 facilities, excluding clinics, was 198, with 63.4% (35/55) having fewer than 200 beds. This proportion is similar to the proportion of hospitals classified by number of beds in Japan described above, suggesting that it could reflect the actual situation of small hospitals in Japan.
3.2. Sequence Types and Carbapenemase Variants in K. aerogenes
Five STs were identified in this study: STs 4, 189, 296, 668, and 709. ST4 is one of the predominant STs in China and has been reported in nosocomial clusters in pediatric hospitals and among men who have sex with men [25,26]. While the international travel history of the patient from whom it was isolated in our study is unknown, the novel detection of ST709, which is the CC of ST4, raises the question of whether this strain will be reported internationally or domestically in the future. However, further studies are needed to confirm this hypothesis. ST189 has been registered in Brazil, but details regarding it are unclear. ST296 was registered in China but was discovered in Brazil before 2016 [27]. Two strains isolated from Osaka belonged to ST668 reported domestically and originated from the same hospital. ST668 has been previously registered in Japan, and given its central position within the CC, it is plausible that ST668 potentially represents one of the prevalent STs in the country. Because of the lack of comprehensive studies on K. aerogenes in Japan, including genetic testing, the mainstream STs are unknown, but some STs of the strains carrying carbapenemase could be clarified. In addition to STs, the type of carbapenemase could be mentioned. The most common carbapenemase gene identified in this study was blaIMP-6, followed by blaIMP-1, blaNDM-1, and blaNDM-5 in order of frequency. According to official reports in 2018, the most prevalent carbapenemase is IMP (85.5%), followed by NDM (10.4%) and KPC (3.4%) [10,12]. The IMP type, which is the most common, is considered to be endemic to Japan. On the other hand, the NDM type was first reported in 2011 and the KPC type in 2014, both of which were classified as carbapenemases of foreign origin [28,29]. Although in this study, the total number of isolates harboring carbapenemase was low, it was noteworthy that relatively more isolates, i.e., 2/6 (33.3%), harbored blaNDM. Owing to the small number of strains, no significant trend was observed in typing with MLST. In addition, no clear trend in the association between the STs and carbapenemase was evident.
3.3. Mobile Genetic Elements in This Study, Including Plasmids, Integrons, and Transposons
Plasmids carrying these carbapenemases and information on integrons and transposons provide valuable insights into the origins of carbapenemase genes [30,31].
As integrons related to blaIMPs in Enterobacterales, two blaIMP-1-containing integrons (In1311 and In1312) have been internationally reported [32]. From China, integron (In1223) has been reported [33]. Moreover, another integron from Enterobacter hormaechei (In1426) and an integron from E. coli (In798) have been reported from Japan [34,35] (Table S3). The structures of these integrons harboring blaIMP-1 are more complex and diverse, varying both nationally and internationally, as well as between different bacterial species, compared with integrons harboring blaIMP-6 (In1321, In1547, and In1690), which are less frequently reported outside of Japan [32,36]. In this study, the integron harboring blaIMP-1 with blaOXA-101 inserted could not be found by BLAST or Integrall. In addition to the integron, TUT0015 showed interesting Inc-type characteristics. IncN4 was previously reported in Italy as a plasmid of Citrobacter freundii carrying blaOXA-181 (GenBank accession number JQ996149). We did not find any reports on the association between the IncN4 plasmid type and blaIMP-1. There is no information available on the plasmids of this Inc type or the integrons they carry, and further research is needed in this area.
As for IMP-6, the epidemic is mostly limited to western Japan [37]. Regarding our three strains harboring blaIMP-6, the integron of the strain isolated in Fukuoka in 2016 (TUT0013) was In1690, which is one of the representative integrons harboring blaIMP-6 and reported in several studies [35,36,38]. The integron (In1547) of TUT0032 was similar to that of In1690, although a transposase (ISVsa10) insertion was observed downstream of aadA2. TUT0023 also lacked the class 1 integrase of In1547, but otherwise, the integron structure was identical. These results suggest that the integrons harboring blaIMP-6 identified in this study are of a type that is relatively common in Japan and could have been transmitted by E. coli or K. pneumoniae, which are more common hosts of blaIMP-6 [39].
In addition to blaIMP carbapenemases, we isolated two bacterial strains that produce blaNDM carbapenemases. For blaNDM, unlike blaIMP, the involvement of transposons in the mobile elements of the resistance genes is important [40]. Recently, two strains of K. aerogenes harboring blaNDM-5 were reported for the first time in Japan [14]. These strains were isolated between 2019 and 2020, which coincided with or slightly followed the period of our isolates. According to the previously mentioned report, when the gene structure was related to blaNDM-5, the 5′ end of the blaNDM-5 sequence in the K. aerogenes NDM-5 plasmid consisted of IS5, ISAba125, IS3000, and Tn2. This structure was very similar to our two blaNDM K. aerogenes strains (TUT0037, TUT0049), although IS5 was missing in the plasmid carrying blaNDM-1 in our case. The plasmids reported in China harboring blaNDM-5 (Genbank accession no. MK450346) and those reported in Japan harboring blaNDM-5 (Genbank accession no. LC54851 and AP019679) also had the same IS element and/or transposase, such as IS5, ISAba125, IS3000, and Tn2 at the 5′ end of blaNDML-5, and both were on IncX3-type plasmids. The structure containing “Tn2-IS3000-ISAba125-IS5” at the 5′ end of blaNDM was suggested to be possibly related to IncX3. Moreover, IS3000 and ISAba125 are considered components of transposons, such as Tn3000 and Tn125. Tn3000 is typically a complex transposon consisting of two copies of IS3000 and is known to contain the structure of Tn125 [41,42]. Although the composite transposon (CT) structure could not be confirmed, the blaNDM-containing structures of TUT0034 and TU0049 suggest that they could be part of the Tn3000 transposon. Tn3000 is common in the genus Klebsiella and is often linked to South Asian origins, especially India, more so than East Asia, with frequently undefined replicon types [40].
This study had several limitations. Owing to the limitations of short-read sequencing, we were unable to elucidate the entire structure of the plasmids, let alone the mobile genetic elements. A more detailed investigation of these structures would require long-read sequencing. Nevertheless, we believe that we were able to discuss the origins of integrons and transposons based on the limited information available. Second, because preserved strains that meet the national criteria for CRE were obtained from among the strains requested for microbiological testing by each medical institution through major testing companies in Japan, detailed patient information (such as medical history, travel history, and outcomes) was not available, and we could not determine whether the isolated strains were the causative agents of infection or merely colonizers.
4. Materials and Methods
4.1. Bacterial Strains
Nonduplicate clinical isolates of carbapenem-resistant K. aerogenes were collected from domestic medical institutions in Japan between September 2014 and December 2022. To avoid the duplication of samples from any single individual, only the first isolate detected in each patient was selected. The identification of bacterial strains and antimicrobial susceptibility test were conducted using Vitek MS (BioMérieux, Marcy l’Étoile, France) and the Microscan WalkAway® system (Beckman Coulter, Brea, CA, USA), with accompanying panels of the Microscan Neg® series (Neg Combo EN 4 J and Neg MIC EN 2 J) (Beckman Coulter, Brea, CA, USA). The bacterial solution was prepared using a prompt inoculation method [43]. Antimicrobial susceptibility was tested according to the guidelines outlined by the Clinical and Laboratory Standards Institute (CLSI) [44,45,46,47]. CRE was defined in accordance with the Japanese Infectious Disease Control Law as Enterobacterales isolates exhibiting a minimum inhibitory concentration (MIC) of ≥2 µg/mL for meropenem, ≥2 µg/mL for imipenem, and ≥64 µg/mL for cefmetazole. Information regarding the size and role of the targeted medical institutions was compiled based on a survey of each hospital’s publicly available hospital profile, number of beds, and bed classification (Table S4).
4.2. Screening of Carbapenemase Genes by PCR
Carbapenemase genes were screened by using the Cica Geneus® Carbapenemase Genotype Detection KIT2 (Kanto Chemical Co., Tokyo, Japan), which can detect the following genes: blaIMP-1, Verona integron-encoded metallo-beta-lactamase (blaVIM), Guiana extended-spectrum (blaGES), blaKPC, blaNDM, blaOXA-48, and blaIMP-6. Bacterial processing and thermal cycling were performed according to the manufacturer’s instructions.
4.3. Whole-Genome Sequencing and Genomic Analysis
DNA was extracted using the QIAamp DNA Mini Kit (QIAGEN, Hilden, Germany), and the library was prepared using the Nextera DNA Flex Library Prep Kit (Illumina, San Diego, CA, USA), employing the Nextera DNA CD Index as the index adapter, as per the manufacturer’s guidelines. Genomic sequencing of K. aerogenes strains positive for carbapenemase genes by PCR was performed on an Illumina iSeq 100 (Illumina, San Diego, CA, USA), utilizing paired-end 150 bp reads.
The sequencing data obtained were annotated using the DNA Data Bank of Japan (DDBJ) Fast Annotation and Submission Tool (DFAST) [48]. Acquired antibiotic resistance genes were identified using ResFinder 4.4.2 (http://genepi.food.dtu.dk/resfinder [accessed on 10 November 2023]) from the Center for Genomic Epidemiology (CGE) [49,50]. Because the blaIMP-1 and the blaIMP-6 differ by only one base (base 640 of 741 bp in length: adenine for blaIMP-1 and guanine for blaIMP-6), the detailed identification of blaIMP-1 and blaIMP-6 was confirmed using the NCBI BLAST database (https://blast.ncbi.nlm.nih.gov/Blast.cgi [accessed on 10 November 2023]) using the nucleotide sequences of their respective representative reference isolates (GenBank accession nos. NG_049172.1 and NG_049220.1), which ultimately led to their identification.
4.4. MLST and Phylogenetic Analysis of Carbapenemase-Producing K. aerogenes
The carbapenemase-producing K. aerogenes isolates were analyzed by the amplification of seven housekeeping genes (dnaA, fusA, gyrB, leuS, pryG, rplB, and rpoB). STs were assigned by querying the Institut Pasteur’s Klebsiella aerogenes PubMLST database (https://pubmlst.org/organisms/klebsiella-aerogenes [accessed on 15 November 2023]). CCs were defined at the single-locus variant level. STs and CCs were assessed using BURST analysis via a PubMLST plugin.
4.5. Analysis of Plasmids, Integrons, and Transposons Associated with Carbapenemase
Inc genes in the plasmids were identified using PlasmidFinder 2.1 (https://cge.food.dtu.dk/services/PlasmidFinder/ [accessed on 10 November 2023]) from the CGE [50,51]. To identify plasmids related to carbapenemase, contigs containing carbapenemase were selectively chosen for analysis. Integron and transposon regions were confirmed using Integron Finder 2.0 and VRprofile2 (https://tool2-mml.sjtu.edu.cn/VRprofile/ [accessed on 7 January 2024]) [52,53]. Specific configurations and sequences of integrons were identified using Integrall (http://integrall.bio.ua.pt/ [accessed on 12 December 2023]) [54]; for configurations and sequences not listed in Integrall, NCBI BLAST was used to search for sequences containing structures around carbapenemase. ISfinder (https://www-is.biotoul.fr/index.php [accessed on 12 December 2023]) and TnCentral (https://tncentral.ncc.unesp.br/ [accessed on 25 January 2024]) were used supplementally to confirm IS elements and transposase, respectively [55,56].
4.6. Data Availability
The WGS data were deposited in GenBank via the DDBJ. BioProject accession number: PRJDB17235. The Biosample numbers for the six strains were as follows: SAMD00670838, SAMD00728333, SAMD00728334, SAMD00728335, SAMD00728336, and SAMD00728337.
4.7. Statistical Analysis
We performed a chi-squared test to check the proportion of the number of carbapenem-resistant K. aereogenes strains in relation to the main function of the hospital and the number of beds. Analysis was performed using JMP Pro 16 statistical analysis software (SAS Institute, 2021, Cary, NC, USA). Differences were considered significant at a corrected p-value < 0.05.
5. Conclusions
Over an 8-year period, we identified 67 carbapenem-resistant K. aerogenes strains from 57 mainly small-to-medium-sized medical facilities across 20 prefectures in Japan, with 9% harboring carbapenemase genes including blaIMP-6, blaIMP-1, blaNDM-1, and blaNDM-5. Because of the small number of strains, no clear trend in the association between the STs and carbapenemase was evident. The gene blaIMP-6 was encoded on the same integron as that reported in K. pneumoniae in Japan, suggesting the possibility of transmission from domestic Enterobacteriaceae. Notably, our study uncovered a potentially novel integron with blaIMP-1 and identified similarities in the transposon structure harboring blaNDM-5 with those previously reported in Japan, suggesting that K. aerogenes harboring blaNDM-5 could unknowingly establish and spread domestically. These findings provide critical insights into the molecular epidemiology of carbapenemase-producing K. aerogenes, underscoring the need for ongoing surveillance and strategic interventions to curb the spread of these resistant pathogens.
Supplementary Materials
The following supporting information can be downloaded from https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/ijms25084494/s1.
Author Contributions
Conceptualization, K.T. and H.K.; data curation and interpretation, K.T. and H.K.; inspection enforcement, R.S. and M.O.; supervision, R.S., M.O. and H.K.; visualization, K.T.; writing of the original draft, K.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Japan Society for the Promotion of Science (JSPS) Grant-in-Aid for Scientific Research (KAKENHI), grant number 21K08504.
Institutional Review Board Statement
This study was approved by the Institutional Review Board of Tohoku University Graduate School of Medicine (IRB numbers: 2018-1-445 and 2022-1-865).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets created and analyzed in the current study are not publicly available because they contain a large amount of detailed patient information. The dataset was owned by the Department of Infectious Diseases, Internal Medicine, Tohoku University Graduate School, and BML Inc., Tokyo, Japan.
Conflicts of Interest
Miho Ogawa and Ryuji Sakata were employed by BML Inc. in Japan. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest. In addition, the authors declare that this study was not funded in any way by private companies and that it was solely funded by the Japan Society for the Promotion of Science (JSPS) Grant-in-Aid for Scientific Research (KAKENHI). The funder had no role in the study design, data collection, analysis, interpretation of data, writing of this article, or decision to submit it for publication.
References
- Álvarez-Marín, R.; Lepe, J.A.; Gasch-Blasi, O.; Rodríguez-Martínez, J.M.; Calvo-Montes, J.; Lara-Contreras, R.; Martín-Gandul, C.; Tubau-Quintano, F.; Cano-García, M.E.; Rodríguez-López, F.; et al. Clinical Characteristics and Outcome of Bacteraemia Caused by Enterobacter cloacae and Klebsiella aerogenes: More Similarities than Differences. J. Glob. Antimicrob. Resist. 2021, 25, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Diene, S.M.; Merhej, V.; Henry, M.; El Filali, A.; Roux, V.; Robert, C.; Azza, S.; Gavory, F.; Barbe, V.; La Scola, B.; et al. The Rhizome of the Multidrug-Resistant Enterobacter aerogenes Genome Reveals How New “Killer Bugs” Are Created Because of a Sympatric Lifestyle. Mol. Biol. Evol. 2013, 30, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Doi, Y.; Bonomo, R.A.; Johnson, J.K.; Simner, P.J.; Antibacterial Resistance Leadership Group. A Primer on AmpC β-Lactamases: Necessary Knowledge for an Increasingly Multidrug-Resistant World. Clin. Infect. Dis. 2019, 69, 1446–1455. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Pagès, J.M. Enterobacter aerogenes and Enterobacter cloacae; Versatile Bacterial Pathogens Confronting Antibiotic Treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Sun, Z.; Zhang, Z. Antimicrobial Resistance of Pathogens Causing Nosocomial Bloodstream Infection in Hubei Province, China, from 2014 to 2016: A Multicenter Retrospective Study. BMC Public Health 2018, 15, 1121. [Google Scholar] [CrossRef] [PubMed]
- Holmes, C.L.; Anderson, M.T.; Mobley, H.L.T.; Bachman, M.A. Pathogenesis of Gram-Negative Bacteremia. Clin. Microbiol. Rev. 2021, 34, e00234-20. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Healthcare-Associated Infections (HAIs). Gram-Negative Bacteria Infections in Healthcare Settings. Available online: https://www.cdc.gov/hai/organisms/gram-negative-bacteria.html (accessed on 20 February 2024).
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef] [PubMed]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215 (Suppl. 1), S28–S36. [Google Scholar] [CrossRef] [PubMed]
- Kamio, K.; Espinoza, J.L. The Predominance of Klebsiella aerogenes among Carbapenem-Resistant Enterobacteriaceae Infections in Japan. Pathogens 2022, 11, 722. [Google Scholar] [CrossRef]
- Oka, K.; Matsumoto, A.; Tetsuka, N.; Morioka, H.; Iguchi, M.; Ishiguro, N.; Nagamori, T.; Takahashi, S.; Saito, N.; Tokuda, K.; et al. Clinical Characteristics and Treatment Outcomes of Carbapenem-Resistant Enterobacterales Infections in Japan. J. Glob. Antimicrob. Resist. 2022, 29, 247–252. [Google Scholar] [CrossRef]
- Surveillance on Carbapenem-Resistant Enterobacteriaceae: Collected in 2018 by National Institute of Infectious Diseases [Japanese]. Available online: https://www.niid.go.jp/niid/ja/cre-m/cre-iasrd/9124-475d01.html (accessed on 15 January 2024).
- Wesevich, A.; Sutton, G.; Ruffin, F.; Park, L.P.; Fouts, D.E.; Fowler, V.G., Jr.; Thaden, J.T. Newly Named Klebsiella aerogenes (Formerly Enterobacter aerogenes) Is Associated with Poor Clinical Outcomes Relative to Other Enterobacter species in Patients with Bloodstream Infection. J. Clin. Microbiol. 2020, 58, e00582-20. [Google Scholar] [CrossRef] [PubMed]
- Kayama, S.; Yu, L.; Kawakami, S.; Yahara, K.; Hisatsune, J.; Yamamoto, M.; Yamamoto, K.; Shimono, N.; Kibe, Y.; Kiyosuke, M.; et al. Emergence of blaNDM-5-Carrying Klebsiella aerogenes in Japan. Microbiol. Spectr. 2022, 10, e0222221. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.; Luterbach, C.L.; Dai, W.; Komarow, L.; Earley, M.; Weston, G.; Herc, E.; Jacob, J.T.; Salata, R.; Wong, D.; et al. Characteristics of Community-Acquired Carbapenem-Resistant Enterobacterales. J. Antimicrob. Chemother. 2022, 77, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Ogawa, M.; Sakata, R.; Kanamori, H. Epidemiological Characteristics of Carbapenem-Resistant Enterobacterales in Japan: A Nationwide Analysis of Data from a Clinical Laboratory Center (2016–2022). Pathogens 2023, 12, 1246. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Deshpande, L.M.; Mendes, R.E.; Doyle, T.B.; Sader, H.S. Prevalence of Carbapenemase Genes Among Carbapenem-Nonsusceptible Enterobacterales Collected in US Hospitals in a Five-Year Period and Activity of Ceftazidime/Avibactam and Comparator Agents. JAC Antimicrob. Resist. 2022, 4, dlac098. [Google Scholar] [CrossRef] [PubMed]
- Rossolini, G.M.; Stone, G.G. Assessment of the In Vitro Activity of Ceftazidime/Avibactam Against a Global Collection of Multidrug-Resistant Klebsiella spp. from the INFORM Surveillance Programme (2015–2017). Int. J. Antimicrob. Agents 2020, 56, 106111. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.Y.; Huang, H.Y.; Zou, H.; Wu, M.L.; Lin, Q.X.; Liu, B.; Huang, S.F. Carbapenem-Resistant Klebsiella aerogenes Clinical Isolates from a Teaching Hospital in Southwestern China: Detailed Molecular Epidemiology, Resistance Determinants, Risk Factors and Clinical Outcomes. Infect. Drug Resist. 2020, 13, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Shi, Q.; Wu, S.; Yin, D.; Peng, M.; Dong, D.; Zheng, Y.; Guo, Y.; Zhang, R.; Hu, F.; et al. Dissemination of Carbapenemases (KPC, NDM, OXA-48, IMP, and VIM) among Carbapenem-Resistant Enterobacteriaceae Isolated from Adult and Children Patients in China. Front. Cell. Infect. Microbiol. 2020, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Surveillance on Carbapenem-Resistant Enterobacteriaceae: Collected in 2019 by National Institute of Infectious Diseases [Japanese]. Available online: https://www.niid.go.jp/niid/ja/cre-m/cre-iasrd/10462-496d01.html (accessed on 15 January 2024).
- Surveillance on Carbapenem-Resistant Enterobacteriaceae: Collected in 2020 by National Institute of Infectious Diseases [Japanese]. Available online: https://www.niid.go.jp/niid/ja/cre-m/cre-iasrd/11520-511d01.html (accessed on 15 January 2024).
- Surveillance on Carbapenem-Resistant Enterobacteriaceae: Collected in 2021 by National Institute of Infectious Diseases [Japanese]. Available online: https://www.niid.go.jp/niid/ja/cre-m/cre-iasrd/12223-522d03.html (accessed on 15 January 2024).
- Tsutsui, A.; Suzuki, S. Japan Nosocomial Infections Surveillance (JANIS): A Model of Sustainable National Antimicrobial Resistance Surveillance Based on Hospital Diagnostic Microbiology Laboratories. BMC Health Serv. Res. 2018, 18, 799. [Google Scholar] [CrossRef]
- Pan, F.; Xu, Q.; Zhang, H. Emergence of NDM-5 Producing Carbapenem-Resistant Klebsiella aerogenes in a Pediatric Hospital in Shanghai, China. Front. Public Health 2021, 9, 621527. [Google Scholar] [CrossRef]
- Cheng, Q.; Ma, Z.; Gong, Z.; Liang, Y.; Guo, J.; Ye, X.; Han, Z.; Yao, Z. Whole-Genome Sequencing Analysis of Klebsiella aerogenes among Men Who Have Sex with Men in Guangzhou, China. Front. Microbiol. 2023, 14, 1102907. [Google Scholar] [CrossRef] [PubMed]
- Morgado, S.; Fonseca, É.; Freitas, F.; Caldart, R.; Vicente, A.C. In-Depth Analysis of Klebsiella aerogenes Resistome, Virulome and Mobilome Worldwide. bioRxiv 2023. bioRxiv:2023.08.15.553288. [Google Scholar] [CrossRef]
- Chihara, S.; Okuzumi, K.; Yamamoto, Y.; Oikawa, S.; Hishinuma, A. First Case of New Delhi Metallo-Beta-Lactamase 1-Producing Escherichia coli Infection in Japan. Clin. Infect. Dis. 2011, 52, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Takahashi, R.; Sawabe, E.; Koyano, S.; Takahashi, Y.; Shima, M.; Ushizawa, H.; Fujie, T.; Tosaka, N.; Kato, Y.; et al. First Report of KPC-2 Carbapenemase-Producing Klebsiella pneumoniae in Japan. Antimicrob. Agents Chemother. 2014, 58, 2961–2963. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Jeong, S.H. Mobile Carbapenemase Genes in Pseudomonas Aeruginosa. Front. Microbiol. 2021, 12, 614058. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Peirano, G.; Bradford, P.A.; Motyl, M.R.; DeVinney, R.; Pitout, J.D.D. Genomic Characterization of IMP and VIM Carbapenemase-Encoding Transferable Plasmids of Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 3034–3038. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yin, Z.; Yin, X.; Fang, H.; Sun, Q.; Tong, Y.; Xu, Y.; Zhang, D.; Feng, J.; Chen, W.; et al. Sequencing of blaIMP-Carrying IncN2 Plasmids, and Comparative Genomics of IncN2 Plasmids Harboring Class 1 Integrons. Front. Cell. Infect. Microbiol. 2017, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Harada, S.; Yahara, K.; Ishii, Y.; Motooka, D.; Nakamura, S.; Akeda, Y.; Iida, T.; Tomono, K.; Iwata, S.; et al. Molecular Characterization of IMP-1-Producing Enterobacter cloacae Complex Isolates in Tokyo. Antimicrob. Agents Chemother. 2018, 62, e02091-17. [Google Scholar] [CrossRef]
- Ikegaya, K.; Aoki, K.; Komori, K.; Ishii, Y.; Tateda, K. Analysis of the Stepwise Acquisition of blaCTX-M-2 and Subsequent Acquisition of Either blaIMP-1 or blaIMP-6 in Highly Conserved IncN-pST5 Plasmids. JAC Antimicrob. Resist. 2023, 5, dlad106. [Google Scholar] [CrossRef]
- Abe, R.; Akeda, Y.; Sakamoto, N.; Kumwenda, G.; Sugawara, Y.; Yamamoto, N.; Kawahara, R.; Tomono, K.; Fujino, Y.; Hamada, S. Genomic Characterisation of a Novel Plasmid Carrying blaIMP-6 of Carbapenem-Resistant Klebsiella pneumoniae Isolated in Osaka, Japan. J. Glob. Antimicrob. Resist. 2020, 21, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, A.; Yahara, K.; Kajihara, T.; Sugai, M.; Shibayama, K. Geographical Distribution of Enterobacterales with a Carbapenemase IMP-6 Phenotype and Its Association with Antimicrobial Use: An Analysis Using Comprehensive National Surveillance Data on Antimicrobial Resistance. PLoS ONE 2020, 15, e0243630. [Google Scholar] [CrossRef]
- Yamagishi, T.; Matsui, M.; Sekizuka, T.; Ito, H.; Fukusumi, M.; Uehira, T.; Tsubokura, M.; Ogawa, Y.; Miyamoto, A.; Nakamori, S.; et al. A prolonged multispecies outbreak of IMP-6 carbapenemase-producing Enterobacterales due to horizontal transmission of the IncN plasmid. Sci. Rep. 2020, 10, 4139. [Google Scholar] [CrossRef]
- Yamamoto, N.; Asada, R.; Kawahara, R.; Hagiya, H.; Akeda, Y.; Shanmugakani, R.K.; Yoshida, H.; Yukawa, S.; Yamamoto, K.; Takayama, Y.; et al. Prevalence of, and risk factors for, carriage of carbapenem-resistant Enterobacteriaceae among hospitalized patients in Japan. J. Hosp. Infect. 2017, 97, 212–217. [Google Scholar] [CrossRef]
- Acman, M.; Wang, R.; van Dorp, L.; Shaw, L.P.; Wang, Q.; Luhmann, N.; Yin, Y.; Sun, S.; Chen, H.; Wang, H.; et al. Role of Mobile Genetic Elements in the Global Dissemination of the Carbapenem Resistance Gene blaNDM. Nat. Commun. 2022, 13, 1131. [Google Scholar] [CrossRef]
- Campos, J.C.; da Silva, M.J.; dos Santos, P.R.; Barros, E.M.; Pereira, O.; Seco, B.M.; Magagnin, C.M.; Leiroz, L.K.; de Oliveira, T.G.; de Faria-Júnior, C.; et al. Characterization of Tn3000, a Transposon Responsible for blaNDM-1 Dissemination Among Enterobacteriaceae in Brazil, Nepal, Morocco, and India. Antimicrob. Agents Chemother. 2015, 59, 7387–7395. [Google Scholar] [CrossRef]
- Ma, L.; Qu, Y.; Wang, W.; Wang, D. Characterization of Klebsiella pneumoniae carrying the blaNDM-1 gene in IncX3 plasmids and the rare In1765 in an IncFIB-IncHI1B plasmid. Front. Cell Infect Microbiol. 2024, 11, 1324846. [Google Scholar] [CrossRef] [PubMed]
- Flejzor, B.; Bokkenheuser, V.D. Performance of the Prompt System in Identification and Antimicrobial Susceptibility Testing of Clinical Isolates. J. Clin. Microbiol. 1985, 21, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; CLSI Supplement, M100-S16; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2006. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; CLSI Supplement, M100-S26; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2015. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Approved Standard M07-A7; CLSI: Wayne, PA, USA, 2006. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Approved Standard M07-A10; CLSI: Wayne, PA, USA, 2016. [Google Scholar]
- Tanizawa, Y.; Fujisawa, T.; Kaminuma, E.; Nakamura, Y.; Arita, M. DFAST and DAGA: Web-Based Integrated Genome Annotation Tools and Resources. Biosci. Microbiota Food Health 2016, 35, 173–184. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Néron, B.; Littner, E.; Haudiquet, M.; Perrin, A.; Cury, J.; Rocha, E.P.C. IntegronFinder 2.0: Identification and Analysis of Integrons Across Bacteria, with a Focus on Antibiotic Resistance in Klebsiella. Microorganisms 2022, 10, 700. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Goh, Y.X.; Tai, C.; Wang, H.; Deng, Z.; Ou, H.Y. VRprofile2: Detection of Antibiotic Resistance-Associated Mobilome in Bacterial Pathogens. Nucleic Acids Res. 2022, 50, W768–W773. [Google Scholar] [CrossRef]
- Moura, A.; Soares, M.; Pereira, C.; Leitão, N.; Henriques, I.; Correia, A. INTEGRALL: A Database and Search Engine for Integrons, Integrases and Gene Cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Ross, K.; Varani, A.M.; Snesrud, E.; Huang, H.; Alvarenga, D.O.; Zhang, J.; Wu, C.; McGann, P.; Chandler, M. TnCentral: A Prokaryotic Transposable Element Database and Web Portal for Transposon Analysis. mBio 2021, 12, e0206021. [Google Scholar] [CrossRef]
Figure 1.
Prefectures and sample counts for collected carbapenem-resistant Klebsiella aerogenes. Listed are the prefectures and the number of clinical isolates (number of facilities). In prefectures where carbapenemase was detected, the type (number of isolates) is noted in bold at the bottom.
Figure 1.
Prefectures and sample counts for collected carbapenem-resistant Klebsiella aerogenes. Listed are the prefectures and the number of clinical isolates (number of facilities). In prefectures where carbapenemase was detected, the type (number of isolates) is noted in bold at the bottom.

Figure 2.
Three clonal complexes identified in this study. Three clonal complexes (CCs) were identified, including ST4, ST668, and ST296. The potential ancestral type (AT) of each CC was ST4, ST668, and ST434. CC1, centered on ST4, was the representative CC in China; ST31, although an AT, was part of the overall CC centered on ST4; ST668 and a CC containing ST296 had not been reported previously.
Figure 2.
Three clonal complexes identified in this study. Three clonal complexes (CCs) were identified, including ST4, ST668, and ST296. The potential ancestral type (AT) of each CC was ST4, ST668, and ST434. CC1, centered on ST4, was the representative CC in China; ST31, although an AT, was part of the overall CC centered on ST4; ST668 and a CC containing ST296 had not been reported previously.

Figure 3.
Linearized genetic environment around blaIMP. The respective integron structure, including the blaIMP, is shown. Dotted lines are nucleotide sequences; major open reading frames (ORFs) in the mobile genetic elements are indicated by blocked arrows. Coding sequences are represented by arrows. The size of the arrow indicates the length of the ORF. An index of 1 kbp is used as a reference for the length. Arrows indicate class 1 integrase (light blue), transposase or IS element (yellow), carbapenemase gene (purple), resistance genes other than carbapenemase (beige), and others (gray). Squares indicate attC.
Figure 3.
Linearized genetic environment around blaIMP. The respective integron structure, including the blaIMP, is shown. Dotted lines are nucleotide sequences; major open reading frames (ORFs) in the mobile genetic elements are indicated by blocked arrows. Coding sequences are represented by arrows. The size of the arrow indicates the length of the ORF. An index of 1 kbp is used as a reference for the length. Arrows indicate class 1 integrase (light blue), transposase or IS element (yellow), carbapenemase gene (purple), resistance genes other than carbapenemase (beige), and others (gray). Squares indicate attC.

Figure 4.
Linearized genetic environment around blaNDM. Gene structures around blaNDM in TUT0037 and TUT0049. Both TUT0037 and TUT0049 had transposases, such as IS3000 and ISAba125 upstream of blaNDM. ISAba125 and IS3000 constituted transposons Tn125 and Tn3000, respectively. Regions considered to be related to Tn3000 are labeled “Tn3000-related regions” and regions considered to be related to Tn125 are labeled “Tn125-related regions”. The orientations and colors of the block arrows are the same as in Figure 3.
Figure 4.
Linearized genetic environment around blaNDM. Gene structures around blaNDM in TUT0037 and TUT0049. Both TUT0037 and TUT0049 had transposases, such as IS3000 and ISAba125 upstream of blaNDM. ISAba125 and IS3000 constituted transposons Tn125 and Tn3000, respectively. Regions considered to be related to Tn3000 are labeled “Tn3000-related regions” and regions considered to be related to Tn125 are labeled “Tn125-related regions”. The orientations and colors of the block arrows are the same as in Figure 3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Basic information on six Klebsiella aerogenes strains harboring carbapenemase, along with genetic testing and antimicrobial susceptibility testing results.
Table 1.
Basic information on six Klebsiella aerogenes strains harboring carbapenemase, along with genetic testing and antimicrobial susceptibility testing results.
| Strain Name | Year of Isolation | Prefecture | Type of Facility | Age (Sex) | Sample | STs | Carbapenemase Gene Type | Replicon Types | Integron and Transposon | Antimicrobial Resistance Genes Except Carbapenemase Gene |
|---|---|---|---|---|---|---|---|---|---|---|
| TUT0013 | 2016 | Fukuoka | Acute | 89 (M) | Bile | 189 | blaIMP-6 | – | In1690 | aadA2, aac(6′)-Ib, sul1, blaSHV-190, oqxA, tet(A), blaCTX-M-2 |
| TUT0015 | 2016 | Fukuoka | Long-term | 87 (F) | Sputum | 4 | blaIMP-1 | IncN4 | Novel class 1 integron | blaOXA-101, aph(3′)-XV, fosA, qnrS1 |
| TUT0023 | 2017 | Osaka | Acute | 67 (M) | Urine | 668 | blaIMP-6 | – | In1547-like | aadA2, aac(6′)-Ib, sul1, fosA7, dfrA14, blaCTX-M-2 |
| TUT0032 | 2018 | Osaka | Acute | 84 (F) | Sputum | 668 | blaIMP-6 | – | In1547 | aadA2, aac(6′)-Ib, sul1, fosA7, dfrA14, blaCTX-M-2 |
| TUT0039 | 2018 | Ishikawa | Acute | 72 (M) | Urine | 709 | blaNDM-5 | IncX3 | Tn3000-like | qnrD1 |
| TUT0049 | 2019 | Kanagawa | Acute | 72 (F) | Vaginal discharge | 296 | blaNDM-1 | IncX3 | Tn3000-like | qnrD1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Takei, K.; Ogawa, M.; Sakata, R.; Kanamori, H. Molecular Epidemiology of Carbapenem-Resistant Klebsiella aerogenes in Japan. Int. J. Mol. Sci. 2024, 25, 4494. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084494
AMA Style
Takei K, Ogawa M, Sakata R, Kanamori H. Molecular Epidemiology of Carbapenem-Resistant Klebsiella aerogenes in Japan. International Journal of Molecular Sciences. 2024; 25(8):4494. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084494
Chicago/Turabian StyleTakei, Kentarou, Miho Ogawa, Ryuji Sakata, and Hajime Kanamori. 2024. "Molecular Epidemiology of Carbapenem-Resistant Klebsiella aerogenes in Japan" International Journal of Molecular Sciences 25, no. 8: 4494. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084494
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.