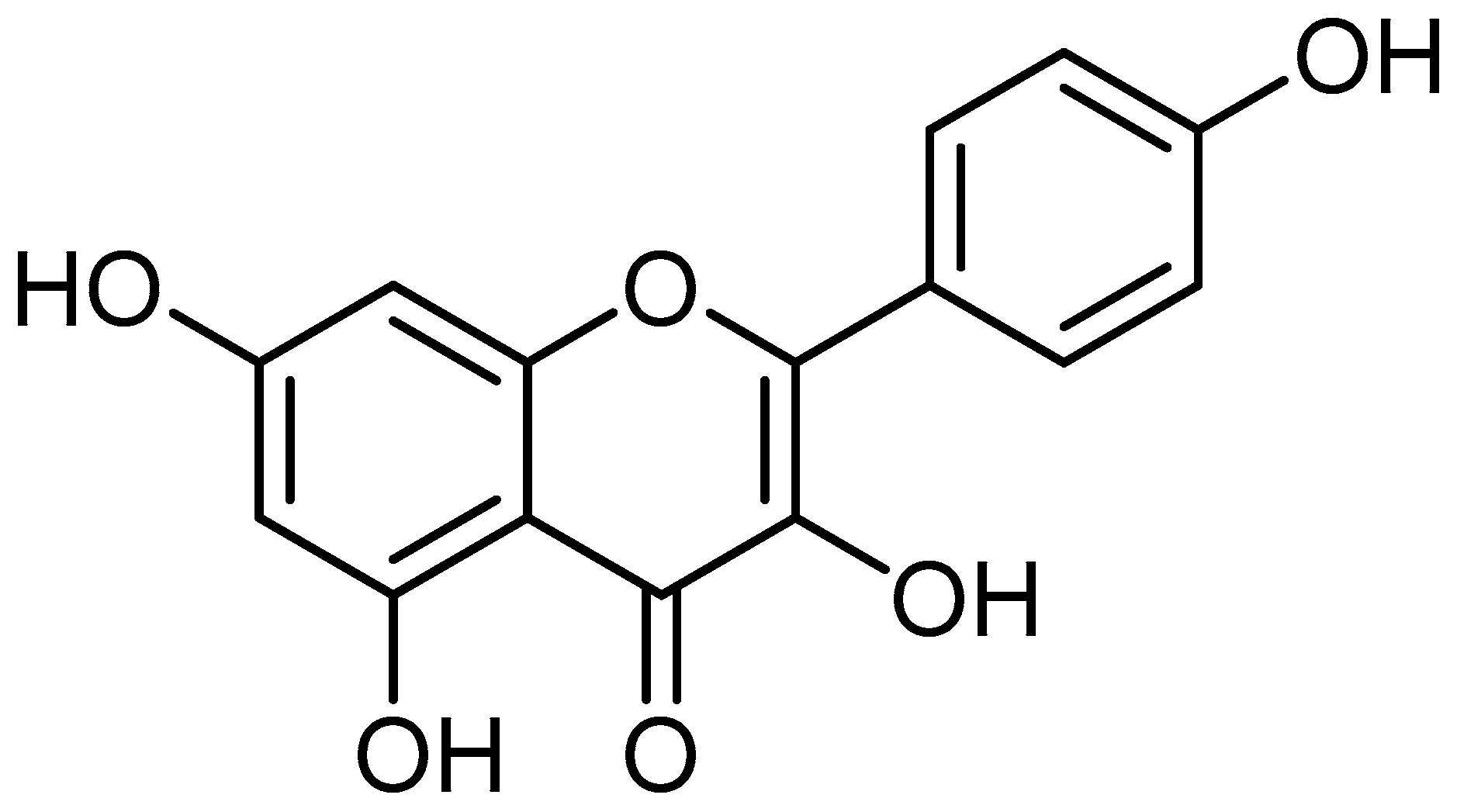
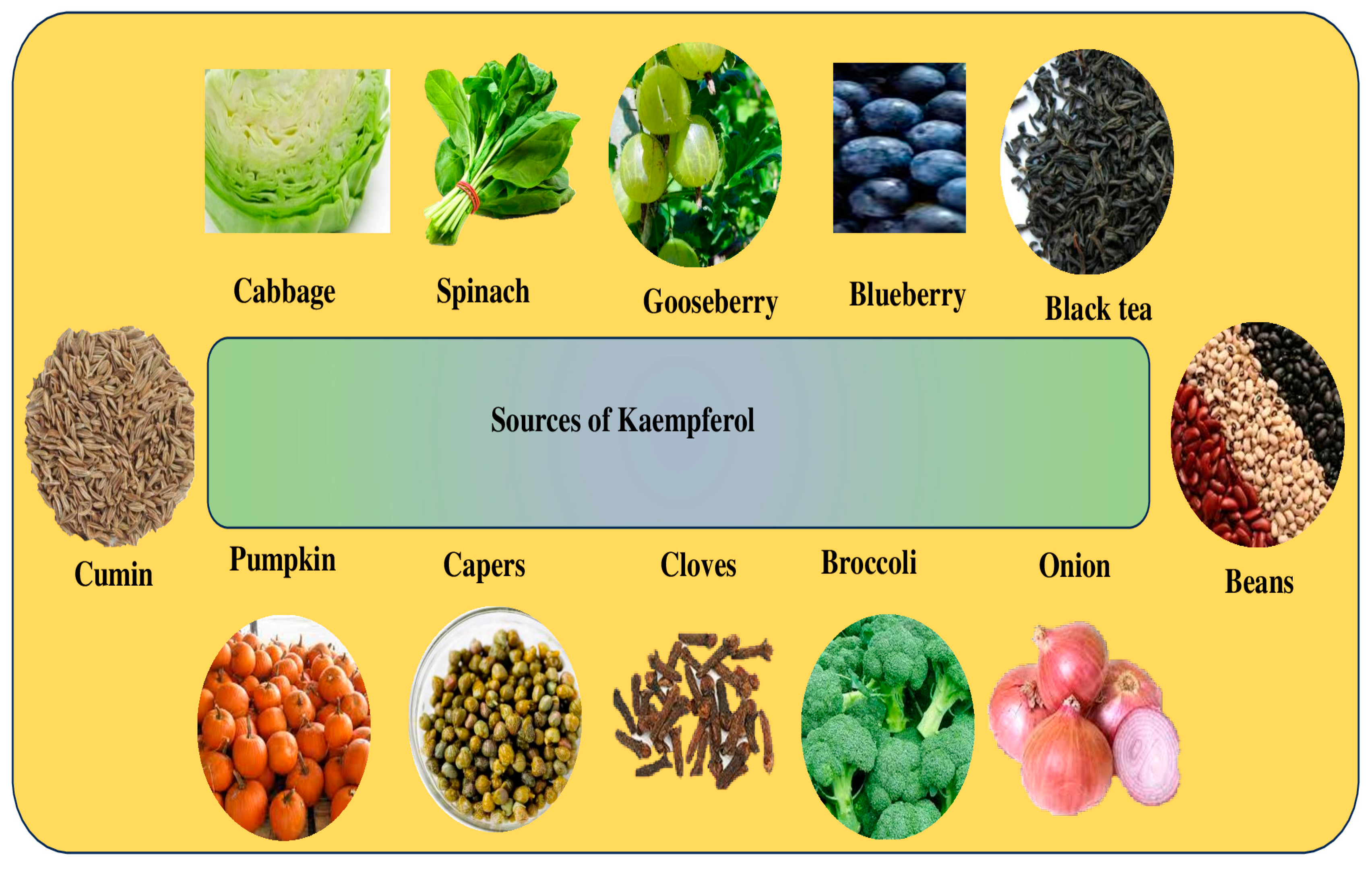
Kaempferol, a flavonoid found in plants, fruits, and spices has various biological activities associated with health, such as anti-oxidant, anti-inflammation, anti-diabetes, neuroprotective, anticancer, and other biological activities. The pharmacological activities of kaempferol in different pathogenesis are described below.
4.1. Antioxidant Potential
Oxidative stress, inflammation, and apoptosis are crucially interlinked processes that hold immense significance in physiological and pathological states [
21]. Oxidative stress occurs when an imbalance exists between producing and accumulating reactive oxygen species (ROS) and antioxidants [
22]. Various natural compounds showed a role in the inhibition of pathogenesis by inhibiting or reducing oxidative stress [
23,
24,
25]. It has been demonstrated that phenolic compounds possess several antioxidant qualities, such as the ability to stop the production of reactive species, scavenge free radicals and neutralize them, form chelate complexes with pro-oxidizing metals, and remove or repair damage caused by reactive chemicals [
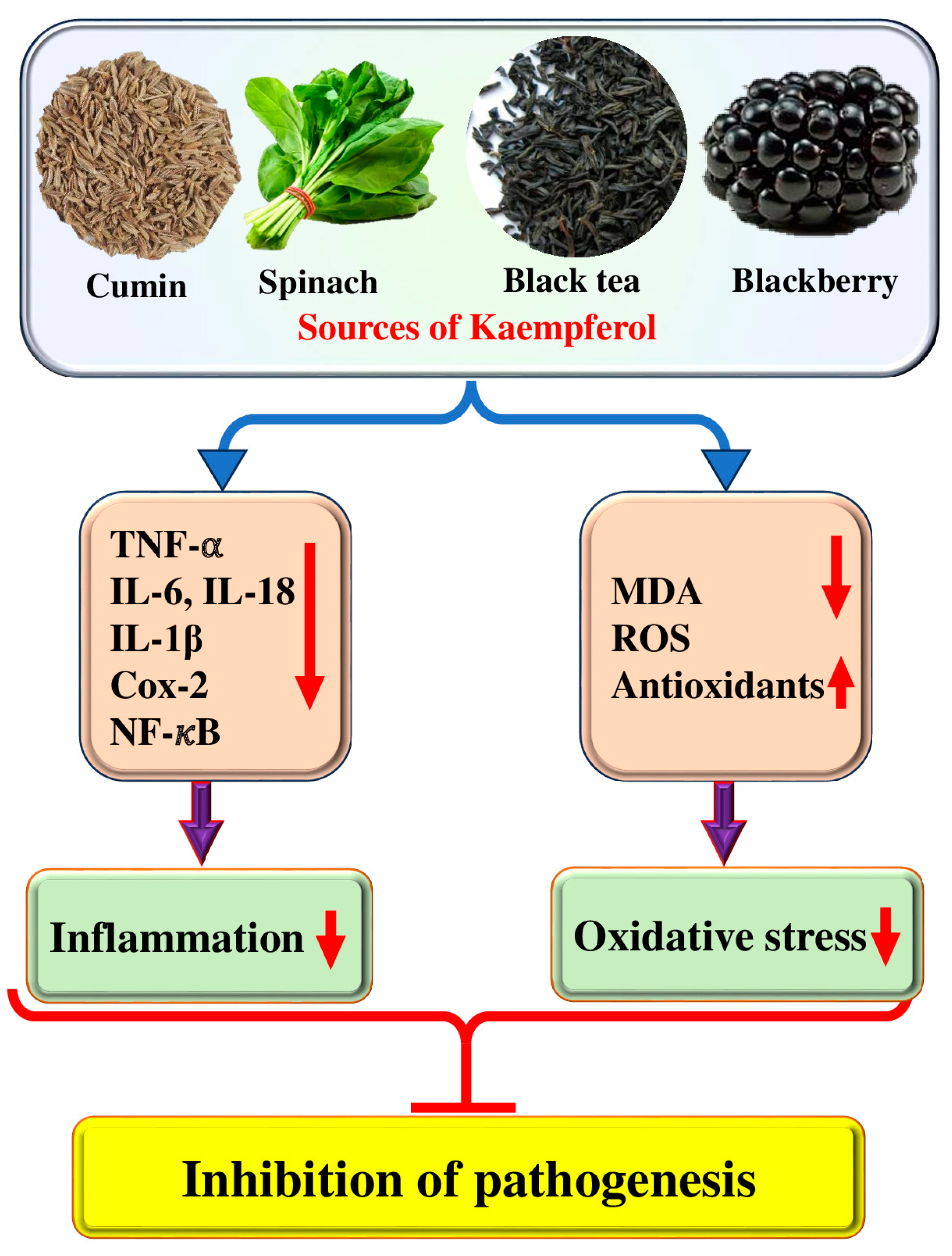
26]. Kaempferol plays a role in disease management through its antioxidant potential and helps cells protect themselves from injury caused by free radicals (
Figure 2).
Kaempferol has been found to protect human retinal pigment epithelium cells (ARPE-19) against the harmful effects of hydrogen peroxide-induced oxidative cell damage and apoptosis. This has been achieved by activating specific signaling pathways involving Bax/Bcl-2 and caspase-3 molecules, as evidenced by real-time PCR and Western blot results. Furthermore, it has been demonstrated that kaempferol suppresses the H
2O
2-induced increase in vascular endothelial growth factor (VEGF) mRNA expression levels in ARPE-19 cells. Additionally, it has been shown to control the activities of superoxide dismutase (SOD) and reactive oxygen species (ROS), which helps maintain the balance of the antioxidant and oxidation systems in ARPE-19 cells exposed to H
2O
2 [
27]. Earlier research has established that administering kaempferol to diabetic rats significantly improved their plasma glucose, insulin, and lipid peroxidation products. The study also found that the levels of enzymatic and non-enzymatic antioxidants were restored to almost normal levels. These findings advise that kaempferol has effective antioxidant properties, which are evident in its ability to increase antioxidant status and decrease lipid peroxidation markers in diabetic rats. This may protect against the risks of diabetic complications [
28].
The amount of oxidative stress was assessed by determining the levels of MDA, ROS, SOD, GSH, and GSH-PX in L2 cells. It was reported that kaempferol has no clear role in ROS, MDA, SOD, and GSH-PX GSH in L2 cells under normal conditions. After suffering A/R, the levels of MDA and ROS in L2 cells were meaningfully increased, while the levels of GSH, SOD, and GSH-PX were meaningfully decreased, proposing that the A/R process produced intense oxidative stress in L2 cells. Moreover, this compound pre-treatment considerably decreased MDA and ROS levels, and increased GSH-PX, SOD, and GSH levels, signifying that kaempferol could efficiently decrease oxidative stress induced by A/R. In vivo investigations have shown that kaempferol is useful in decreasing pathological damage, preventing oxidative stress and apoptosis, elevating the expressions of mitochondrial cytochrome c and Bcl-2, and lowering those of cytoplasmic cytochrome c and Bax in the lung tissues of rats following I/R. These results imply that kaempferol may be helpful in shielding lung tissues from the damaging effects of I/R [
29]. A study was performed to examine whether kaempferol affects oxidative stress and inflammation in the heart, liver, and lungs after hemorrhagic shock. The SOD activities and MDA levels in the liver, heart, and lung were measured to examine the role of kaempferol on oxidative response following hemorrhagic shock (HS). It was found that MDA levels were meaningfully higher in the HS groups as compared to the Sham group, whereas SOD activities were diminished in the HS groups as compared to the Sham group. The injection of kaempferol following HS exhibited no effects on MDA levels in the heart, liver, and lung, and on SOD activities in the lung heart, and liver compared with the HS group. However, the kaempferol injection 12 h earlier to the induction of HS caused increased SOD activities and reduced MDA levels as compared with the HS group in the lung, heart, and liver. The pretreatment of hemorrhagic shock mice with kaempferol has shown a significant reduction in plasma levels of TNF-α and IL-6, as well as a reversal of MPO, SOD, and MDA in the heart, lung, and liver. Additionally, an increase in the expression of HO-1 was observed in these same organs. These findings suggest that kaempferol could potentially be beneficial in mitigating the adverse effects of hemorrhagic shock in mice [
30].
4.2. Anti-Inflammatory Potential
The release of reactive species by inflammatory cells during inflammation can lead to oxidative stress. This highlights the close connection between oxidative stress and inflammation [
31]. Natural products and their compounds have been discovered to possess protective properties against inflammation [
32] and kaempferol’s role as an anti-inflammatory has been confirmed (
Figure 2 and
Table 1). Dilip Sharma et al., 2019, reported that kaempferol inhibits hyperglycemia-induced activation of RhoA and decreases oxidative stress and fibrosis (extracellular matrix protein and TGF-β1 expression) in RPTEC and NRK-52E cells. TNF-α and IL-1β levels were noticeably increased in high glucose treated cells as compared to normal glucose state. This compound caused a dose-dependent decrease in TNF-α and IL-1β levels in cells [
33]. Moreover, at concentrations of 12.5 and 25 μg/mL, kaempferol has been shown to significantly suppress the release of IL-1β, TNF-α, IL-18, and IL-6 while also inhibiting the activation of NF-κB and Akt in lipopolysaccharide plus ATP-induced cardiac fibroblasts. The findings suggest that kaempferol reduces inflammation in cardiac fibroblasts by suppressing the activation of Akt and NF-κB [
34]. Another study finding reported that kaempferol reduced IL-32-induced monocyte differentiation to product macrophage-like cells. Furthermore, kaempferol has been shown to inhibit the activation of p38 induced by nuclear factor-κB and IL-32 in a dose-dependent manner in THP-1 cells. Kaempferol has also been found to improve the production of inflammatory mediators such as TSLP, TNF-α, IL-1β, IL-8, and nitric oxide, induced by lipopolysaccharide, in macrophage-like cells differentiated by IL-32 [
35]. The study revealed that the oral administration of kaempferol at a 50 mg/kg dose significantly inhibited the antigen-induced passive cutaneous anaphylaxis response in IgE-sensitized mice [
36]. Model rabbits fed with a high-cholesterol diet developed noteworthy progression of atherosclerosis. Compared to the control group, the serum levels of blood lipids, IL-1β, TNF-α, and MDA were significantly elevated, while SOD levels were reduced in the model rabbits. The gene and protein expressions of VCAM-1, E-sel, ICAM-1, and MCP-1 in atherosclerotic aortas showed a significant increase in the model group. However, in comparison to the model rabbits, the levels of IL-1β, TNF-α, and MDA decreased significantly, while serum SOD activity increased. Additionally, the gene and protein expressions of VCAM-1, MCP-1, E-sel, and ICAM-1 in the aortas showed a significant decrease with the administration of kaempferol [
11]. Kaempferol demonstrates the anti-inflammatory effect through suppressing the translocation of CagA as well as VacA proteins and leading to pro-inflammatory cytokines down-regulation [
37].
It has been reported in earlier studies that kaempferol has the capability to hinder the growth of both unstimulated and IL-1β-stimulated RASFs. Additionally, it can also lower the production of MMP-1, COX-2, MMP-3, and PGE2, which are induced by IL-1β. Additionally, it has been reported that kaempferol can inhibit the activation of NF-κB and the phosphorylation of ERK-1/2, p38, and JNK. These proteins are known to play a role in the inflammatory response in RA. Based on these findings, it has been suggested that kaempferol may have the potential as a therapeutic agent for the treatment of RA [
38]. It has been reported that at doses ranging from 1 to 20 μmol/L, kaempferol has a dose-dependent effect on the reduction in TNFα-induced expression of epithelial intracellular cell adhesion molecule-1. Additionally, kaempferol has been found to inhibit eosinophil integrin β2 expression, which may hinder the eosinophil–airway epithelium interaction. It has also been reported that kaempferol has the ability to reduce TNFα-induced airway inflammation by decreasing monocyte chemoattractant protein-1 transcription. Moreover, oral administration of kaempferol decreased OVA challenge-elevated expression of eotaxin-1 and eosinophil, major essential proteins, through the blockade of NF-κB transactivation [
39]. Kaempferol has shown promising results in increasing the expression of FOXP3, a protein-coding gene expressed in Treg cells, and amplifying mRNA levels of FOXP3 and IL-10 in Treg cells. This suggests that kaempferol may have the potential as a treatment option for autoimmune diseases [
40].
Table 1.
Anti-inflammatory potential of kaempferol with different mechanisms. The downward pointing arrow indicates decrease.
Table 1.
Anti-inflammatory potential of kaempferol with different mechanisms. The downward pointing arrow indicates decrease.
| Study Types | Doses | Mechanism | Outcome of the Study | Refs. |
|---|
| In vitro | 10, 50 Μm | TNF-α & IL-1β ↓ | | [33] |
| In vivo | 12.5 and 25 μg/mL | TNF-α, IL-1β, IL-6, and IL-18 ↓ | | [34] |
| In vitro | 0.02, 0.2, and 2 mg/mL | IL-1b, TNF-a, and IL-8 ↓ | | [35] |
| In vivo | 150 and 30 mg/kg | TNF-α, IL-1β, ICAM-1, VCAM-1 ↓ | | [11] |
| In vivo and in vitro | 10 or 20 mg/kg
1–20 mmol/L | NF-kB ↓ | | [39] |
| In vitro | 40 μM | NF-κB ↓ | | [41] |
| In vitro | 5 to 200 μmol/L | iNOS, COX-2 ↓ | It was noticed that at all concentrations, kaempferol reduced the levels of iNOS, COX-2, and CRP protein
| [42] |
4.3. Anti-Diabetic Potential
Diabetes is a chronic metabolic disorder that affects a large population worldwide. The current mode of treatment is effective but it also causes adverse effects. In this regard, natural compounds have proven their role as anti-diabetic through the modulation of various biological activities. It is interesting to note that natural compounds such as kaempferol have been found to have anti-diabetic properties (
Table 2). The anti-diabetic potential of kaempferol has been evidenced by the modulation of various biological activities (
Figure 3). The anti-diabetic potential of kaempferol was examined, and it was reported that both fasting and non-fasting blood glucose levels of diabetic mice were decreased by kaempferol treatment. Further, STZ-induced diabetic mice exhibited severe glucose intolerance, which was improved by the treatment of kaempferol. Kaempferol therapy gradually increased the activity of hexokinase in the liver and skeletal muscle of diabetic mice, while decreasing the activity of gluconeogenesis and hepatic pyruvate carboxylase [
43]. The study assessed kaempferol’s protective role against oxidative stress in streptozotocin (STZ)-induced diabetic rats. It was noticed that diabetic rats presented a slowly raised level of plasma glucose on the initial, 7th, 15th, and 45th days, correspondingly, and a reduced insulin level on the 45th day as compared to normal group rats. Kaempferol oral administration or glibenclamide in diabetic rats exhibited a reduced level of plasma glucose as well as an increased insulin level as compared to diabetic control rats. Moreover, lipid peroxidation levels exhibited a noteworthy increase in diabetic rats, while, upon treatment with kaempferol or glibenclamide, these lipid peroxidative markers displayed a substantial decline towards a normal level. The administration of kaempferol or glibenclamide significantly increased the activities of SOD, CAT, GPx, and GST, which had decreased in the tissues of diabetic rats [
28]. It was discovered that giving obese mice oral kaempferol at a dose of 50 mg/kg/day—the human equivalent of 240 mg/day for an average 60 kg human—markedly improved blood control. This activity was related to the reduction in hepatic glucose production in addition to improved whole-body insulin sensitivity, without altering body weight gain, food consumption, or adiposity. The same study found that kaempferol treatment enhanced hexokinase besides Akt activity while decreasing pyruvate carboxylase in addition to glucose-6 phosphatase activity in the liver of obese mice [
44]. Kaempferol has been shown to improve insulin sensitivity and blood lipid levels dose-dependently. It has also been found to restore insulin resistance and induce changes in glucose disposal. Moreover, kaempferol inhibited the IkB kinase α (IKKα) and IkB kinase β (IKKβ) and phosphorylation of insulin receptor substrate-1 (IRS-1) [
45]. A study assessed kaempferol’s ability to prevent diabetic nephropathy caused by streptozotocin. Renal levels of TNF-α and IL-6, cleaved caspase-3, p38, and Bax were reduced by Kaempferol. This suppressed JNK phosphorylation as well as NF-κB p65 transactivation, along with upregulation of Bcl-2. In both control and STZ-diabetic rats, Kaempferol reduced fasting glucose levels, increased fasting insulin levels and HOMA-β, stimulated SOD and GSH levels, reduced the levels of ROS and MDA, and increased the expression of Nrf2 and HO-1 [
46].
4.6. Renoprotective Effects
Kidney-associated pathogenesis affects a large population worldwide and is a significant cause of morbidity and mortality. The current mode of treatment causes adverse effects in various ways. In this regard, natural compounds have proven their role as renoprotective through the modulation of various biological activities. Natural compounds have been shown to have renoprotective properties (
Table 3). Kidney diseases have become a significant public health concern worldwide due to their association with severe clinical complications [
63,
64,
65]. It is well-established that oxidative stress and inflammation play substantial roles in the development and progression of chronic kidney disease (CKD) [
66]. Renal fibrosis, a hallmark of CKD, is characterized by various factors, such as the accumulation of inflammatory cells, injury to renal tubules, and the development of tubulointerstitial fibrosis [
67]. It is crucial to address this issue to ensure better health outcomes for individuals affected by such conditions. Kidney disease is a very concerning health issue that, unfortunately, leads to high morbidity and mortality rates even with the availability of advanced diagnostic and management tools [
64,
65,
68]. It is essential to look into more effective ways to prevent and treat the progression of acute kidney injury and chronic kidney disease [
64,
65]. There is a need to explore different therapeutic strategies that can help manage these conditions effectively. The renoprotective potential of kaempferol has been evidenced by the modulation of various biological activities (
Figure 3).
The nephroprotective effectiveness of kaempferol and apigenin as dietary supplements in cisplatin-induced renal injury was examined. Findings revealed from MTT assay data, morphology studies, comet, and ROS analysis suggest that CIS 11.36 μM + kaempferol 25 μg/mL and CIS 11.36 μM + apigenin 12.5 μg/mL protect against cisplatin-induced nephrotoxicity. The finding of western blot analysis additionally advises the involvement of NGAL in the apigenin and kaempferol-mediated nephroprotection [
69]. Studies have shown that pre-treatment with kaempferol can help reduce the oxidative stress, inflammation, and apoptosis caused by cisplatin. This can lead to a betterment in renal injury and overall kidney functioning. In addition to reducing oxidative stress, inflammation, and apoptosis caused by cisplatin, studies have found that pre-treatment with kaempferol also led to a decrease in cisplatin-induced phosphorylation of p38, JNK, and ERK1/2 in renal tissues [
70]. The role of kaempferol in the regulation of blood glucose levels was measured by GLP-1 as well as insulin release during OGTT in C57BL/6 mice. It was noticed that the administration of kaempferol (100 and 200 mg/kg) caused a dose-dependent enhancement in the release of GLP-1. This event was noticeable by a progressive rise in the levels of GLP-1 up to 60 min of treatment, but then again, the highest levels of GLP-1 release were attained 45 min post-administration of kaempferol. Moreover, the insulin levels were meaningfully decreased 19417 by 4.1-fold in diabetic nephropathy (DN) mice when compared to the control group. After 10 days of kaempferol treatment, insignificant changes in GLP-1 as well as insulin levels were noticed. However, after 21 days of treatment, Kaempferol (200 mg/kg) meaningfully increased the GLP-1 as well as insulin levels by 1.5-fold. Kaempferol has shown the potential to improve renal fibrosis and histological changes. It also reduces the expression of DN markers such as CTGF, TGF-β1, fibronectin, collagen IV, RhoA, IL-1β, ROCK2, and p-MYPT1 in DN kidney tissues. An increase in the expression of nephrin and E-cadherin was also found in the same study [
47].
4.7. Neuroprotective Effects
Neurological disease is a significant cause of morbidity and mortality. The current mode of treatment causes adverse effects. In this regard, natural compounds have proven their role as neuroprotective through the modulation of various biological activities such as inflammation and oxidative stress.
Oxidative stress and inflammation can play essential roles in developing various diseases and conditions. Chronic inflammation in the central nervous system has been widely studied and identified as a possible factor in the development and progression of various neurodegenerative diseases [
71]. Activated microglia, the immune cells that dwell within the central nervous system, have been recognized as a major factor in the development of neuroinflammation [
72]; oxidative stress is a significant factor in aging [
73], and it is a universal mechanism causing cell death [
74]. Natural compounds such as kaempferol have been shown to have anti-diabetic properties (
Table 3). Studies have shown that kaempferol can enhance the activity and levels of antioxidant enzymes like superoxide dismutase (SOD) and glutathione (GSH) [
75]. To measure the role of kaempferol, rat pheochromocytoma cells (PC12) and mice were used as neuronal models. In vitro assay-based findings described that kaempferol was revealed to have a protective role against oxidative stress-induced cytotoxicity in PC12 cells. Kaempferol administration significantly upturned amyloid beta peptide (Abeta)-induced impaired performance [
76]. ICV streptozotocin (3 mg/kg) administration was performed on the first and the third day of the surgery, and the animals’ memory was examined via passive avoidance tasks. The ICV injections of streptozotocin meaningfully decreased memory retention as well as intact pyramidal cells as compared to the control group. The kaempferol improved the effects of streptozotocin. Results demonstrated that kaempferol can optimize cognitive deficits caused by streptozotocin injections and also has some valuable effects on hippocampal CA1 pyramidal neurons [
77]. A study was conducted to evaluate the effects of kaempferol on various factors such as cognitive impairment, lipid peroxidation, hippocampal antioxidants, neuro-inflammation markers, and apoptosis in rat models of sporadic Alzheimer’s disease. The results exhibited a substantial improvement in memory and spatial learning as demonstrated by shortened escape latency as well as searching distance in the Morris water maze in the ovariectomized + streptozotocin + kaempferol group compared with the ovariectomized + streptozotocin group. Kaempferol also showed important increases in brain levels of antioxidant enzymes of glutathione and superoxide dismutase, with a reduction in malondialdehyde and tumor necrosis factor-α. This finding reveals that kaempferol is capable of improving streptozotocin-induced memory impairment in ovariectomized rats, possibly via elevating endogenous hippocampal antioxidants of glutathione superoxide and dismutase and decreasing neuroinflammation. This study advises that kaempferol may be a possible neuroprotective agent against cognitive deficits in Alzheimer’s disease [
78]. The research findings indicate that when Alzheimer’s disease flies were exposed to kaempferol, it delayed the loss of their climbing ability and memory. Additionally, kaempferol reduced the activity of acetylcholinesterase and oxidative stress [
79].
The study focused on evaluating the neuroprotective effects of kaempferol on hippocampal neuronal cells (HT22) that were exposed to glutamate. The experiment yielded interesting results. It was found that the administration of kaempferol (25 μM) had a significant positive effect on cell viability compared to the control group. Additionally, the neuroprotective properties of kaempferol are believed to be due to its ability to regulate the expression levels of various proteins, including AIF (apoptosis-inducing factor), Bcl-2, Bid, and mitogen-activated protein kinase. Based on these findings, it is suggested that kaempferol could be a promising candidate for pharmacological interventions aimed at both preventing and treating neurodegenerative diseases, such as Alzheimer’s disease [
80].
Table 3.
Role of kaempferol in the management of liver, kidney, and nerve-associated pathogenesis.
Table 3.
Role of kaempferol in the management of liver, kidney, and nerve-associated pathogenesis.
| Activity | Doses | Outcome of the Study | Refs. |
| Hepatoprotective | 250 g/kg, orally | This compound reduced liver damage, inflammatory markers, and liver function enzymes. Increased the antioxidant enzymes | [53] |
| 10, 20.0 mg/kg | It is worth noting that kaempferol is a powerful compound that can decrease oxidative stress and lipid peroxidation while simultaneously enhancing antioxidative defense activity. | [54] |
| 5 and 10 mg/kg | Kaempferol hepatic histology and serum parameters and reduced the levels of pro-inflammatory mediators. | [55] |
| 4.9 mg/kg | Kaempferol reduced TBARS and TNF-α levels | [57] |
| 250 and 500 mg/kg | Kaempferol inhibited the synthesis of collagen and activation of hepatic stellate cells | [58] |
| 30 and 60 mg/kg | This compound had a positive effect on liver health. Specifically, this compound was found to decrease the levels of alanine aminotransferase and aspartate aminotransferase | [60] |
| 20 mg/kg/day | Kaempferol may decrease the expression level of LXRα and LPCAT3, thus improving inflammation | [60] |
| 125 mg/kg | Kaempferol injury through anti-inflammatory, anti-oxidative, as well as anti-apoptotic activities | [61] |
| Gastroprotective | 40, 80, or 160 mg/kg | This compound decreased the ulcer index, completely protected the mucosa from lesions, and decreased pro-inflammatory cytokine activity. | [62] |
| Nephroprotective | 100 and 200 mg/kg | Kaempferol reduced inflammation, oxidative stress, and apoptosis as well as bettered renal injury and its functioning | [70] |
| 100 and 200 mg/kg | Kaempferol maintains histological changes as well as renal fibrosis | [47] |
| Neuroprotective | 10 mg/kg | Kaempferol treatment increases enhanced acquisition as well as retrieval of spatial memory | [75] |
| 10 mg/kg | Kaempferol increases antioxidant enzymes whereas it reduces malondialdehyde and tumor necrosis factor-α | [78] |
| 0, 20, 30, and 40 μM | Kaempferol reduced the acetylcholinesterase and oxidative stress activity and delayed the loss of climbing ability and memory | [79] |
4.8. Cardioprotective Activity
Cardiovascular disease, such as acute myocardial infarction, has unfortunately become a significant risk factor for human health [
81]. Natural compounds have been shown to play a significant role in the management of cardiovascular disease. Many of these compounds are found in numerous foods and supplements. Consumption of these natural compounds in diet can help reduce the risk of cardiovascular disease and improve overall heart health. Kaempferol has been shown to have cardioprotective properties (
Table 4). A critical study evaluated the kaempferol on cardiac hypertrophy and the underlying mechanism. Kaempferol has been shown to reduce cardiac hypertrophy caused by aorta banding significantly. This is evidenced by decreased cardiomyocyte areas and interstitial fibrosis, reduced apoptosis, and improved cardiac functions. It has been observed in in vitro experiments that kaempferol has the ability to prevent the activity of the ASK1/JNK1/2/p38 signaling pathway, as well as the enlargement of H9c2 cardiomyocytes [
82]. Kaempferol pretreatment (1–10 mg/kg i.p. before DOX administration) showed a dose-dependent recovery of heart and body weights. DOX also caused oxidative stress damage, as indicated by decreased catalase and superoxide dismutase activities in rat hearts and increased lactate dehydrogenase activity in serum. All these actions were dose-dependently bettered by kaempferol, demonstrating its efficiency in counteracting DOX-induced oxidative stress. In addition, in vitro studies have suggested that kaempferol may have mitochondrion-dependent pathways to counteract the cardiotoxicity caused by DOX. It has been observed that this counteraction is achieved through the inhibition of p53 expression and its binding to the promoter region of the Bax proapoptotic gene, while not affecting the Bcl-2 antiapoptotic gene in vitro [
83]. Another study reported that kaempferol mitigated hypertrophy and cardiac dysfunction caused by cisplatin. Moreover, pretreatment with kaempferol decreased cisplatin-induced cardiomyocyte apoptosis.
Moreover, cisplatin decreased the expression of BCL-2 and increased the expression of BAX, but such changes initiated by cisplatin were overturned by the treatment of kaempferol. In H9c2 cells, kaempferol intensely reduced cisplatin-induced apoptosis as well as inflammatory response by modulating the STING/NF-κB pathway [
84]. Kaempferol pretreatment has been shown to improve the recovery of LVDP and ±dp/dt max, while also increasing the levels of SOD, the GSH/GSSG ratio, and P-GSK-3β. In addition, it has been observed that kaempferol pretreatment led to a decrease in myocardial infarct size, along with decreased levels of cytoplasmic cytochrome C, creatine kinase (CK), cleaved caspase-3, lactate dehydrogenase (LDH), tumor necrosis factor-alpha (TNF-α), and malondialdehyde (MDA) [
85].
Table 4.
The cardioprotective activity of kaempferol.
Table 4.
The cardioprotective activity of kaempferol.
| Types of Study | Model | Dose | Outcome the Study | Refs. |
|---|
| In vivo | Mice subjected to aorta banding | 100 mg/kg/d | Kaempferol meaningfully reduced cardiac hypertrophy caused by aorta banding Decreased cardiomyocyte areas as well as interstitial fibrosis, Decreased apoptosis and improved cardiac functions.
| [82] |
| In vitro | H9c2 cardiomyocytes
stimulated by phenylephrine | 25 μM | | [82] |
| In vivo | DOX-induced cardiotoxicity | 1–20 mg/kg | | [83] |
| In vitro | DOX-induced cardiotoxicity | 0 to μ50 M | | [83] |
| In vivo | Cisplatin-induced cardiac injury | 10 mg/kg | | [84] |
| In vitro | H9c2 cells model | 1, 5, or 10 Μm | Kaempferol showed a role in the inhibition of cisplatin-caused cell death Treatment with kaempferol decrease HMGB1, MCP-1, IL-6 and TNF-α
| [84] |
| In vivo | Myocardial Ischemia/Reperfusion Injury | 15 mmol/L | | [85] |
| In vitro | H/R-induced injury | 5, 10, 20, or 30 μM | Kaempferol reduces H/R-induced the damages Down-regulation of lactate dehydrogenase (LDH) activity, the increases in anti-apoptotic protein
| [86] |
| In vivo | 5-Fluorouracil-Induced Cardiotoxicity | 1 mg/kg | | [87] |
| In vivo | ISO-induced cardiac injury | 3 and 10 mg/kg | Kaempferol decrease in serum CK-MB, LDH, troponin-I and lipid profile MDA level and MMP-2 expression and MMP-9 level was reduced by kaempferol
| [88] |
| In vivo | Ischemia-Reperfusion Injury | 20 mg/kg | | [89] |
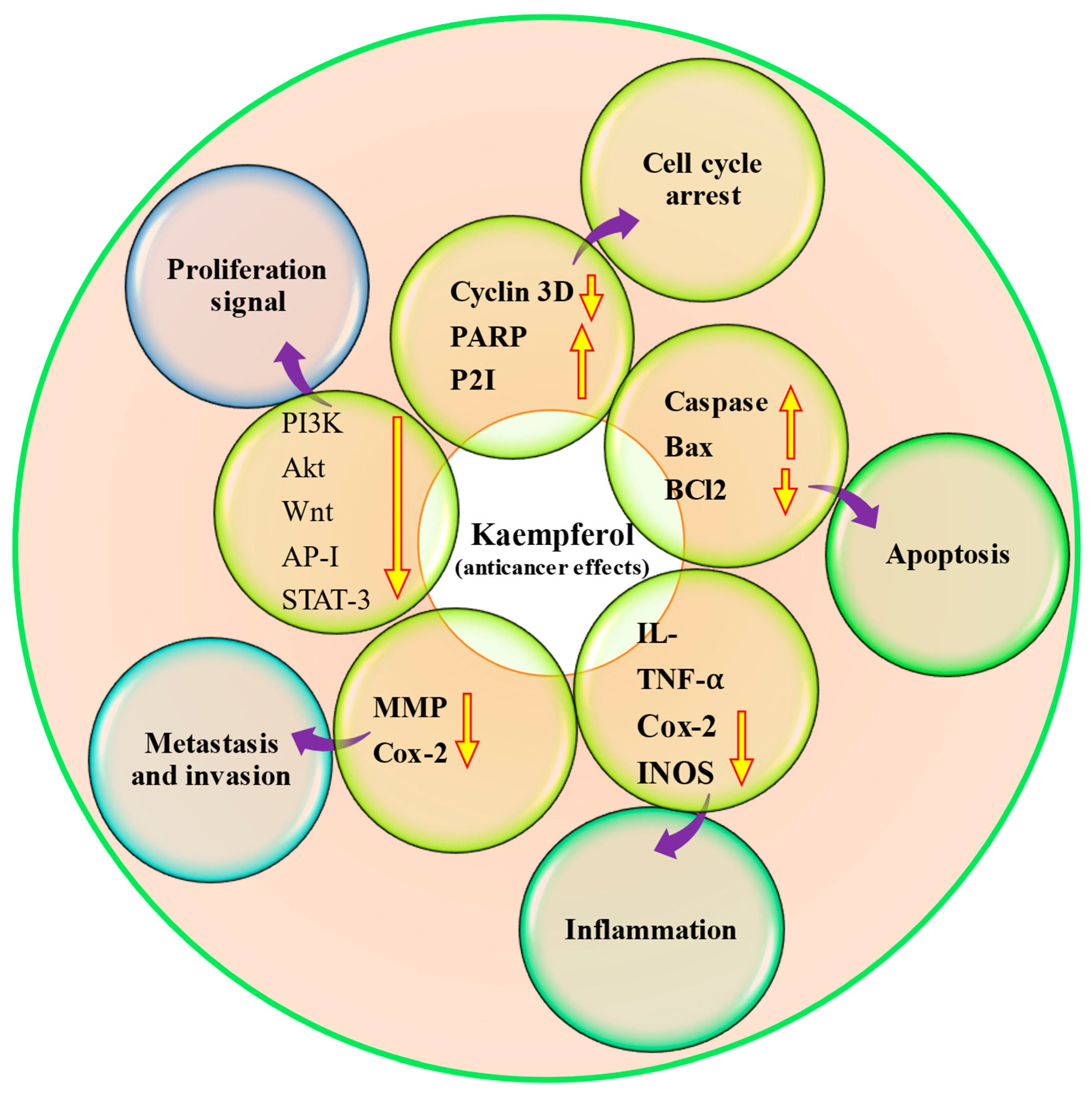
4.9. Anti-Cancer Potential
Cancer is a multifactorial disease, and unfortunately, it is one of the significant causes of mortality worldwide. It is unfortunate that despite the development of various treatment strategies, cancer remains a substantial cause of death worldwide [
90,
91]. Cancer treatment modules such as chemotherapy drugs, like doxorubicin, cisplatin, and fluorouracil, and surgical removal are commonly used to treat most cancers. Still, they can also lead to severe side effects and toxicity. Interestingly, many therapeutic drugs used today have their roots in natural resources like alkaloids, taxanes, and flavonoids [
92]. There has been a growing interest in using natural compounds in chemotherapy to enhance the effectiveness of anti-neoplastic drugs [
93] and inhibit cancer growth through the modulation of cell signaling molecules [
94,
95,
96,
97,
98]. Kaempferol has confirmed the anti-cancer role by modulating different cell signaling molecules (
Table 5 and
Figure 4). According to a breast cancer study, kaempferol decreased the expression of IQGAP3 in breast cancer cells. In these cancer cells, kaempferol effectively inhibited proliferation and promoted death while decreasing IQGAP3 expression. It was noticed that upregulating IQGAP3 expression prevented cancer cells from undergoing apoptosis. This was correlated with a rise in the expression of B cell lymphoma 2 (Bcl2) and phosphorylated extracellular signal-regulated kinases 1/2 (p-ERK1/2), as well as a fall in the expression of Bcl-2-associated X protein (Bax). However, this effect was counteracted by the treatment of kaempferol [
99]. Furthermore, kaempferol showed a role in the inhibition of MCF-7 breast cancer cell growth, possibly by downregulating Bcl2 expression and inducing apoptosis. Kaempferol is a hopeful cancer preventive as well as therapeutic agent for breast cancer [
100]. A recent study based on ovarian cancer reported that kaempferol plays a role in the management of ovarian cancer through the regulation of the cell cycle as kaempferol-induced G2/M cell cycle arrest through the Chk2/p21/Cdc2 pathway and Chk2/Cdc25C/Cdc2 pathway. Further, after the treatment with 40 μM kaempferol, the viability of ovarian cancer cells was reduced to 59% kaempferol-induced apoptosis in cancer cells. The late apoptotic rate of A2780/CP70 cells was meaningfully increased to 23.95% when treated with kaempferol (40 μM) for 48 h [
101]. Kaempferol has the ability to inhibit the migration, adhesion, and invasion of human breast carcinoma cells (MDA-MB-231). In addition, it was observed that kaempferol led to a decrease in the activity and expression of MMP-2 and MMP-9 [
102]. Kaempferol works synergistically with cisplatin in preventing cell viability of ovarian cancer, and their inhibition on cell viability was brought through inhibiting ABCC6 and cMyc gene transcription. Results reported as cisplatin treatment, with or without kaempferol, did not alter mRNA levels meaningfully for ABCC1, NFκB1, and ABCC5, genes. However, cisplatin reduced cMyc and ABCC6 mRNA levels in a dose-dependent way, with a remaining mRNA level of around 58% at a 80 μM concentration. Kaempferol treatment inhibited cMyc and ABCC6 gene transcription down to 68%. The combination of kaempferol and 40 μM cisplatin prevented ABCC6 as well as cMyc genes mRNA levels down to 65%, and 80 μM cisplatin leads to the lowest mRNA level of 55% for cMyc, though no enhancement is noticed for ABCC6 gene at 80 μM cisplatin concentration. The combination of the two chemicals takes the CDKN1A mRNA level up to 1064.7%. Moreover, the apoptosis assay displayed that the addition of kaempferol (20 μM) to the cisplatin treatment induces the apoptosis of the cancer cells [
103]. Moreover, kaempferol time-dependently prevented VEGF secretion and decreased in vitro angiogenesis. Kaempferol decreased ERK phosphorylation and NFκB and cMyc expression. In contrast, it promoted p21 expression. To explore whether ERK signaling participated in kaempferol’s regulation of VEGF, it transfected ovarian cancer cells with ERK1 plasmid and noticed that kaempferol’s inhibition on VEGF transcriptional activation, as reflected by VEGF reporter luminescence, was stopped by forced ERK1 expression in a concentration-dependent way, and a significance was attained in A2780/CP70 cells. This finding proposed that not only does this flavonoid inhibit ERK phosphorylation as well as VEGF expression in ovarian cancer cells, but this prevention of VEGF is also dependent on, at least partially, ERK signaling suppression [
104]. It was observed that kaempferol treatment led to time-dependent increases in PTEN expression in bladder cancer cells, especially in those treated with 40 μM kaempferol. Notably, treatment with 40 μM kaempferol resulted in a substantial decrease in the expression of Ser473-phosphorylated Akt. Taken together, these results indicate that kaempferol has the ability to upregulate PTEN expression and prevent pAkt (Ser473) in EJ cells. [
105].
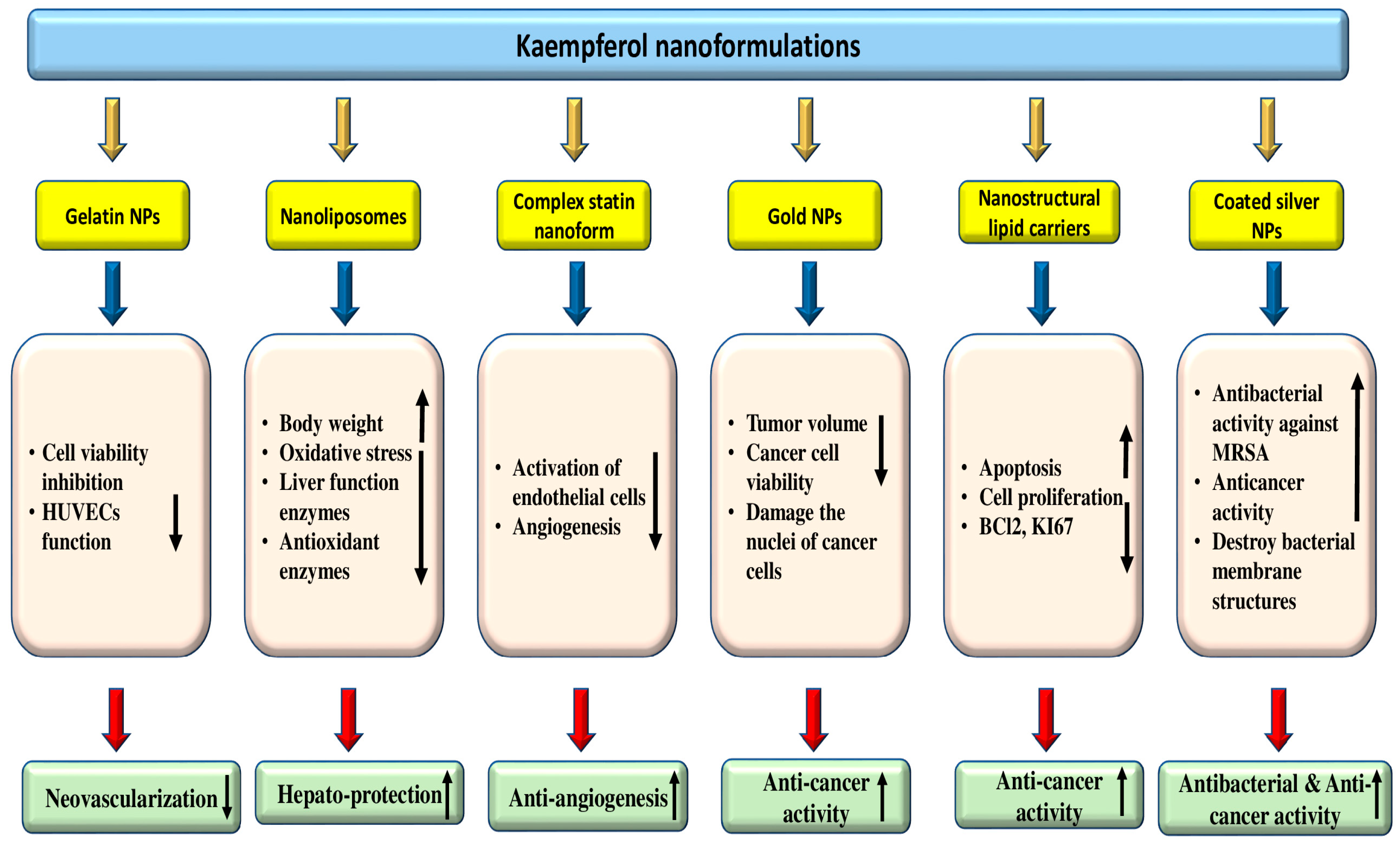
4.10. Anti-Microbial Effects
The issue of antibiotic resistance, particularly in Gram-negative strains, is quite alarming, especially in hospitals where vulnerable patients are at a higher risk [
106]. However, it is not just the hospitals affected by multidrug-resistant (MDR) strains; our food supply also poses a threat due to the rampant use of antibiotics in livestock for infection treatment, growth promotion, and prevention [
107]. The extracts from various parts of plants, including roots, stems, fruits, and flowers, are often used to inhibit the growth of microorganisms [
108]. It is worth noting that in addition to their antimicrobial activities, it has the potential to be used as preservatives [
109,
110]. Kaempferol has some promising antimicrobial properties that could potentially be useful in preventing the development and progression of diseases (
Table 6 and
Figure 5).
Some of the anti-microbial effects are described here as:
It has been discovered that kaempferol inhibits the action of bacterial efflux pumps and
S. aureus PriA helicase (SaPriA), which can effectively stop antibiotic-resistant
S. aureus. This has been shown to increase the effectiveness of antimicrobial treatment [
111,
112]. Another study result reported that kaempferol showed antibacterial activities against Propionibacterium acnes. Kaempferol 3-
O-β-(2″-acetyl)-galactopyranoside joined with quercetin showed substantial antibacterial activity through the apoptosis pathway, and it is reported that kaempferol 3-
O-β-(2″-acetyl)-galactopyranoside was found in clusiacea [
113]. The mechanism causing the antimicrobial effect of the antibacterial activity compounds from
H. ascyron L. was examined. The quercetin and kaempferol 3-
O-β-(2″-acetyl)-galactopyranoside were noticed from fraction 8 by means of mass spectrometry as well as nuclear magnetic resonance. It was demonstrated that kaempferol 3-
O-β-(2″-acetyl)-galactopyranoside coupled with quercetin showed noteworthy antibacterial activity through the apoptosis pathway, and it is reported that kaempferol 3-
O-β-(2″-acetyl)-galactopyranoside was found in clusiacea [
114]. The antibacterial activity tests of four flavonoid derivatives, scandenone (1), tiliroside (2), quercetin-3,7-
O-alpha-L-dirhamnoside (3), and kaempferol-3,7-
O-alpha-L-dirhamnoside (4), are presented. It has been reported that all the compounds tested exhibit considerable activity against
S. aureus and
E. faecalis with MIC values of 0.5 microg/mL. The compounds also show moderate inhibition against
K. pneumoniae (4 microg/mL),
E. coli (2 microg/mL),
A. baumannii (8 microg/mL), and
B. subtilis (8 microg/mL) [
115]. The extract of
B. chinense, which holds kaempferol-3-
O-β-
d-rutinoside and kaempferol, showed effectiveness against this bacterium [
116]. A small molecule called kaempferol has anti-biofilm action and specifically prevents
S. aureus biofilms from forming. Using fluorescence microscopy and crystal violet (CV) staining, kaempferol (64 μg/mL) prevented biofilm formation by 80%. Kaempferol inhibited the expression of adhesion-related genes, the activity of
S. aureus sortase (SrtA), and the primary attachment phase of biofilm formation. These results suggest that kaempferol provides a foundation for developing a new anti-biofilm drug that could lower bacterial drug resistance and prevent infections caused by biofilms caused by
S. aureus [
117]. When azithromycin and kaempferol are taken together, as compared to when they are taken alone, ARSA-induced osteomyelitis in rats is decreased by oxidative stress reduction, SAPK and ERK1/2 phosphorylation inhibition, and biofilm formation inhibition [
118].
- II.
Anti-fungal activities
Fungal infections are a cause for concern as they can lead to serious health problems and even life-threatening diseases. They can manifest as acute or chronic illnesses, such as allergic bronchopulmonary aspergillosis or less severe infections like
Candida vaginitis or oral candidiasis [
119]. Dermatophytosis is a fungal infection that touches a significant portion of the population, with an estimated 20–25% of people affected by this condition. It can be a particularly severe type of fungal infection that can cause damage to tissues, organs, and nerves [
120]. Numerous natural compounds or bioactive compounds show a role in anti-fungal activity. Kaempferol 3-
O-b-
d-kaempferol 3-
O-b-
d-glucopyranoside and kaempferol-3-
O-[3-
O-acetyl-6-
O-(E)-p-coumaroyl]-b-d-glucopyranoside isolated from
S. hymettia was noticed to be active against
C. tropicalis,
C. albicans, as well as C.
glabrata [
121]. The study focused on determining kaempferol’s minimum inhibitory concentration (MIC) against both the planktonic and biofilm forms of the
Candida parapsilosis complex. It was observed that the MIC ranges for kaempferol were 32–128 μg ml
−1. Furthermore, kaempferol demonstrated efficacy in reducing the metabolic activity and biomass of developing
C. parapsilosis complex biofilms [
122]. Kaempferol-(coumaroyl glucosyl)-rhamnoside, present in the Trachyspermum ammi extract, has been shown to potentially inhibit the growth of
Candida spp. [
123].
- III.
Anti-viral activities
Viral infections have been a persistent threat to human health worldwide. Viruses are involved in various types of pathogenesis, including cancer. The vast range of natural products presented delivers an auspicious avenue for the discovery of new and powerful antiviral drugs. Another study based in vitro was performed to examine the antiviral effects of kaempferol against a varicella-zoster virus (VZV) clinical isolate. It was noticed that kaempferol meaningfully inhibited VZV replication without showing cytotoxicity. This compound caused its antiviral potential at the same stage of the VZV life cycle as acyclovir, which prevents VZV DNA replication. Overall, this finding proposes that kaempferol inhibits VZV infection by hindering the DNA replication stage in the viral life cycle [
124]. According to the study, kaempferol has been found to inhibit viral penetration and replication stages, resulting in a significant decrease in virus load by 4-fold and 30-fold, respectively. The study also suggests that kaempferol can effectively inhibit virus replication if added within 16 h post-infection (HPI), reducing the number of DNA copies [
125]. It is becoming more and more essential to develop effective antiviral treatments. The study found that administering kaempferol at a dose of 15 mg/kg via the intragastric route reduced symptoms associated with H9N2 influenza virus infection in BALB/C mice. In particular, myeloperoxidase activity, pulmonary capillary permeability, lung wet/dry weight, inflammatory cell count, and pulmonary edema were all decreased by kaempferol in the mice.
Additionally, it was noticed that kaempferol increased superoxide dismutase activity while decreasing ROS activity, malondialdehyde formation, and the synthesis of TNF-α, IL-1β, and IL-6 [
126]. The study indicated that administering kaempferol at 100 μmol/L completely inhibited the replication of bovine herpesvirus 1 in bovine kidney cells. Furthermore, it was found that kaempferol affects viral replication at the post-entry stages. This suggests that kaempferol has potent antiviral properties, which could be attributed to its inhibition of protein kinase B (Akt) signaling [
127]. The study evaluated the effectiveness of kaempferol, kaempferol glycosides, and acylated kaempferol glucoside derivatives in blocking the 3a channel. The results suggested that viral ion channels could be a good target for developing antiviral agents. The study also found that kaempferol glycosides were effective candidates for inhibiting 3a channel proteins of coronaviruses [
128].
A recent study described flavonoids as promising natural drugs for the treatment of human herpesviruses (HHVs) infections of the nervous system including alpha-herpesviruses, beta-herpesviruses, and gamma-herpesviruses [
129]. Moreover, berberine, an alkaloid, and its role in the inhibition of different pathogenesis, including anti-viral, has been proven. An important study presented the anti-human oncogenic herpesviruses (HOHVs) properties of berberine and mechanisms and pathways induced by this alkaloid through targeting the herpesvirus life cycle as well as the pathogenesis of the linked malignancies [
130].
Another study based in vitro reported kaempferol to have vigorous antiviral activity against bovine herpesvirus 1 (BoHV-1) replication. Kaempferol (100 μmol/L concentration) inhibited viral replication in MDBK cells. It chiefly affects the viral replication at the post-entry stages. Moreover, at a concentration of 25 and 50 μmol/L, kaempferol could meaningfully decrease the expression of inflammatory mediators such as macrophage inflammatory protein 1 alpha, tumor necrosis factor-alpha (TNF-α), and interleukin-8 in human promonocytic U937 cell-derived macrophages (dU937) in response to LPS stimulation [
127]. In a study, flavonols quercetin 3-
O-rutinoside and kaempferol 3-
O-rutinoside derived from Lespedeza bicolor were found to combat HSV-1. They were able to block viral infection by inhibiting viral DNA replication and also exhibited virucidal effects [
131]. The antiviral effects of kaempferol against a varicella-zoster virus (VZV) clinical isolate were investigated. It was noticed that kaempferol meaningfully prevented VZV replication without showing cytotoxicity. This flavonoid employed its antiviral effect at a similar stage of the VZV life cycle as acyclovir, which prevents VZV DNA replication. Overall, this finding proposes that this flavonoid inhibits VZV infection by blocking the stage of DNA replication in the viral life cycle [
124]. A recent study demonstrates the involved mechanisms by which flavonoids decode their antiviral potentials against herpes simplex virus (HSV). Flavonoids are formidable contenders in the battle against HSV infections as they disrupt key stages of the viral life cycle, such as attachment to host cells, entry, DNA replication, latency, and reactivation [
132].
- IV.
Anti-parasitic action
The goal of the study is to determine how kaempferol interacts with hamster neutrophils and impacts
E. histolytica trophozoites. The hamster model is used to evaluate the susceptibility. The results of the investigation showed that trophozoites’ amoebic vitality was dramatically decreased after 90 min of incubation with 150 μM kaempferol. Kaempferol is an effective drug against
E. histolytica through the decrease in expression of
E. histolytica’s antioxidant enzymes. Moreover, studies have shown that kaempferol controls a number of neutrophil processes, such as myeloperoxidase (MPO), reactive oxygen species (ROS), and nitric oxide (NO) [
133]. Kaempferol is capable of inducing the appearance of perinuclear and periplasmic spaces that are devoid of cytosolic content, as well as multilamellar structures. Moreover, it has been discovered that kaempferol causes proapoptotic death, which is linked to a partial arrest in the S phase of the cell cycle. On G. duodenalis trophozoites, kaempferol has been observed to have a proapoptotic impact. This entails a partial stop of DNA synthesis without oxidative stress or damage to chromatin or cytoskeletal components [
134]. The number of adult and encysted larvae was much lower in the groups that received a combination of drugs. In addition, the thickness of the larvae’s capsular layer decreased and the inflammation of the muscles and intestines significantly improved. When combined with albendazole, kaempferol has demonstrated promise as an anti-trichinellosis drug by reducing inflammation and larval capsule formation [
135].
Table 6.
The anti-microbial potential of kaempferol.
Table 6.
The anti-microbial potential of kaempferol.
| Activity | Doses | Outcome of the Study | Refs. |
|---|
| Antibacterial | 0, 30, 60 μg | | [113] |
| Antibiofilm | 64 μg/mL | | [116] |
| 1 μg/mL and 1 mg/mL | | [117] |
| Anti-fungal | 0.25 to 256 mg ml−1 | | [122] |
| | 5, 7, 10, and 15 μg/mL | | [124] |
| 52.40 Μm | | [125] |
| Anti-parasitic | 150 μM | | [129] |
| 20 mg/kg | | [136] |
4.17. Role of Respiratory System
Respiratory disease including asthma, bronchitis, COPD, and ARDS affects a large number of populations worldwide and are a significant cause of morbidity and mortality. However, the current mode of treatment can cause adverse effects. In this respect, natural compounds or their bioactive ingredients have proven to be effective in inhibiting respiratory system-associated pathogenesis by modulating various biological activities.
A study on allergic asthma reported that, in the TGF-β1-induced human bronchial epithelial cells (BEAS-2B), the NOX4 expression was reduced with a kaempferol dose increase. The secretions of IL-33 and IL-25 and the NOX4-mediated autophagy were meaningfully decreased by treatment of kaempferol in the TGF-β1-induced BEAS-2B. Furthermore, in the OVA-challenged mice, kaempferol treatment showed a role in the improvement of airway inflammation as well as remodeling via suppressing NOX4-mediated autophagy. This study has found that kaempferol binds NOX4 to perform its functions in the treatment of allergic asthma, which could potentially provide an effective therapeutic strategy for the further treatment of asthma [
164]. Human airway epithelial BEAS-2B cells and eosinophils were used in the study to examine how kaempferol affected airway inflammation linked to endotoxins or cytokines. The suppression of LPS-induced eotaxin-1 protein production by kaempferol (1–20 μmol/L) may have been caused by Janus kinase 2 (JAK2) JAK2 signaling. The study also found that kaempferol, at doses ranging from 1 to 20 μmol/L, could dose-dependently reduce TNFα-induced expression of epithelial intracellular cell adhesion molecule-1 as well as eosinophil integrin β2, which hindered the eosinophil–airway epithelium interaction. Moreover, oral administration of kaempferol was observed to reduce OVA challenge-raised expression of eotaxin-1 and eosinophil major basic proteins [
39]. A study was performed to explore the ant-ischemia-reperfusion injury (LIRI) mechanism of kaempferol. The study showed that pre-treatment with kaempferol significantly improved the mitochondrial membrane potential, increased cell viability, increased the expressions of Bcl-2 and mitochondrial cytochrome, inhibited the opening of mitochondrial permeability transition pores, reduced the expressions of Bax in addition to cytoplasmic cytochrome c, and reduced the levels of oxidative stress and apoptosis in L2 cells after A/R insult. In rats’ lung tissues after I/R, kaempferol improved pathological damage, raised the expressions of Bcl-2 in addition to mitochondrial cytochrome c, reduced the expressions of Bax besides cytoplasmic cytochrome c, and suppressed the levels of oxidative stress after apoptosis [
29]. Kaempferol pretreated mice presented an important reduction in water content in the lungs. This compound pretreatment displayed a decrease in cytokines IL-1β, IL-6, and TNF-α in lung tissue as well as plasma as a comparison to septic mice without pretreatment. Nonenzymatic antioxidant GSH activities and antioxidant enzymes SOD and catalase were increased with the pretreatment of kaempferol in septic mice. Additionally, kaempferol pretreatment reduced the lung tissue nitrite level besides the iNOS level in septic mice. An important decrease in mRNA expression of iNOS and ICAM-1 was noticed with this pretreatment. Mice pretreated with kaempferol followed by sepsis exhibited more arranged alveolar structures and reduced infiltration of cells in histopathological analysis [
165]. Oral administration of kaempferol (≤20 mg/kg) obstructed bovine serum albumin (BSA) inhalation-induced epithelial cell excrescence as well as smooth muscle hypertrophy through reducing the induction of COX2 and the formation of PGF2α and PGD2, together with decreasing the anti-α-smooth muscle actin expression in mouse airways. This compound prevented the antigen-induced mast cell activation of cytosolic phospholipase A2 responsive to protein kinase Cμ besides extracellular signal-regulated kinase. Additionally, the antigen-challenged activation of the Syk-phospholipase Cγ pathway was diminished in these flavonoid-supplemented mast cells [
166].

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








