
Implications of the Propagation Method for the Phytochemistry of Nepeta cataria L. throughout a Growing Season

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Nepetalactone Concentrations
2.1.1. Pittstown
2.1.2. Upper Deerfield
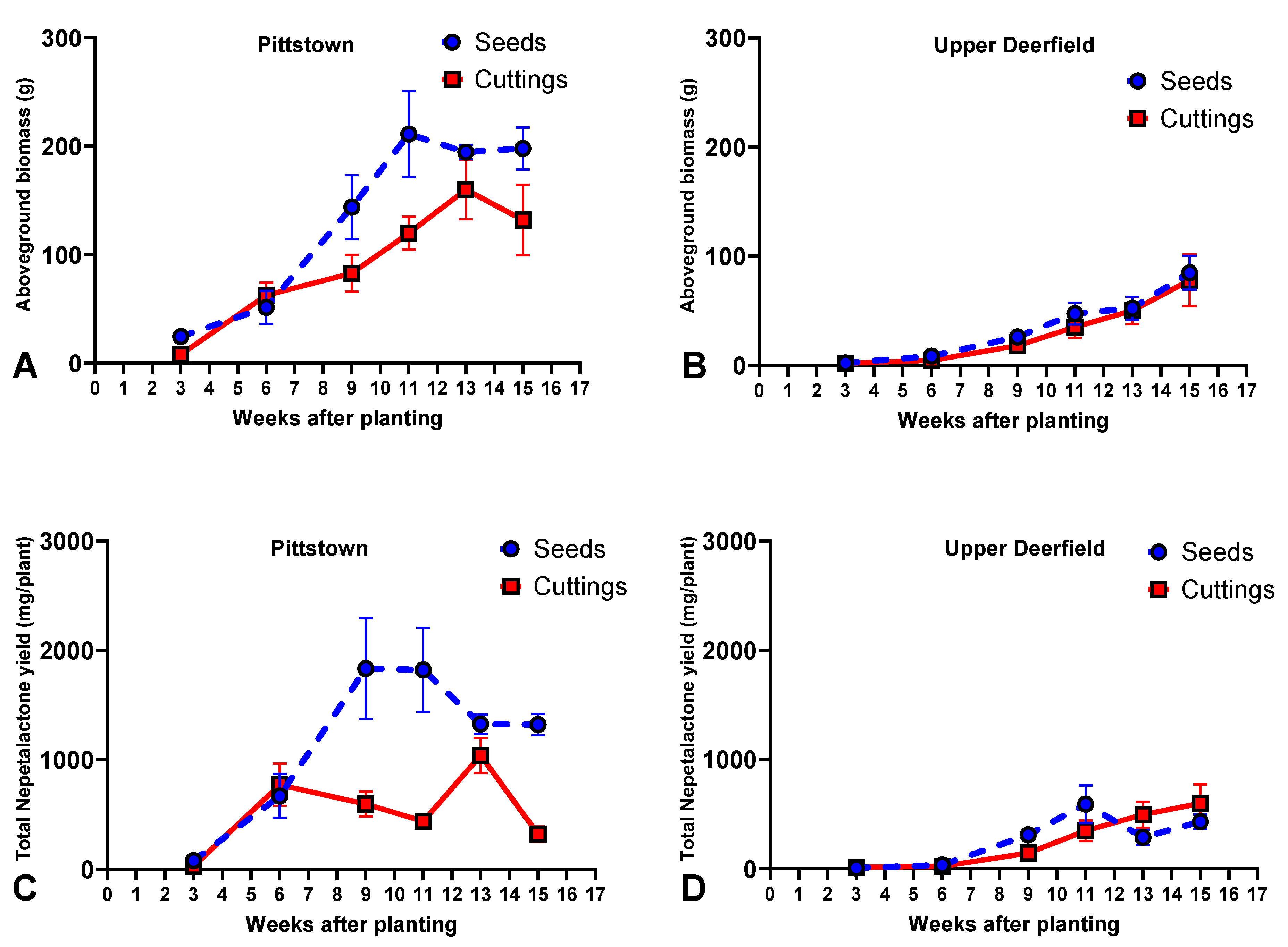
2.2. Biomass Accumulation and Total Nepetalactone Yield
2.2.1. Pittstown
2.2.2. Upper Deerfield
2.3. Nepetalic Acid, Dihydronepetalactone and Nepetalactam Concentrations
2.3.1. Pittstown
2.3.2. Upper Deerfield
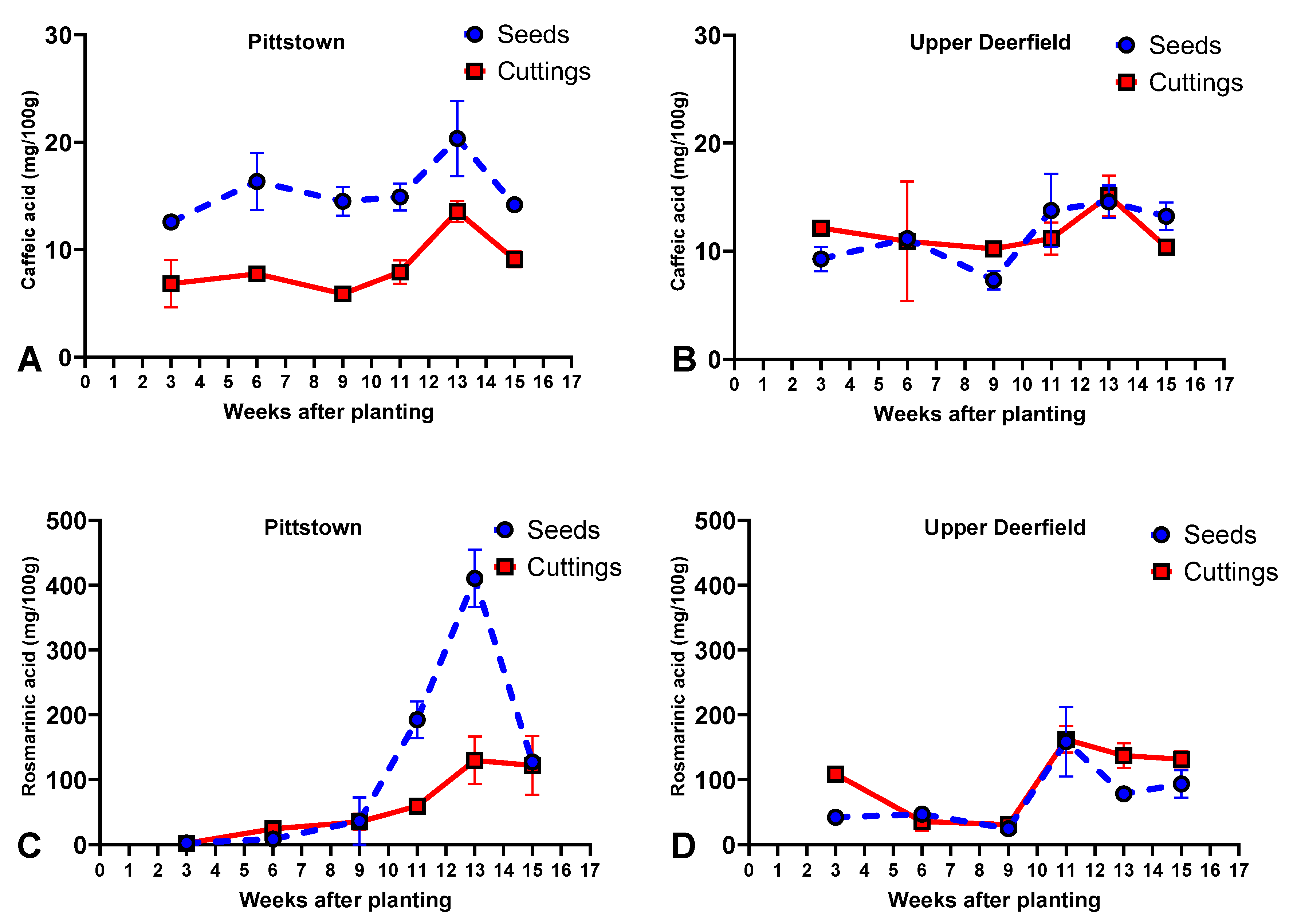
2.4. Caffeic Acid and Rosmarinic Acid
2.4.1. Pittstown
2.4.2. Upper Deerfield
2.5. Apigenin, Luteolin, and Flavone Glycosides
2.5.1. Pittstown
2.5.2. Upper Deerfield
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Month | Ac Prec * | Avg RH | Avg Soil Temp 10 cm | Avg Temp | Max Temp | Min Temp | Avg SolarRad |
|---|---|---|---|---|---|---|---|---|
| mm | % | °C | W/m2 | |||||
| Pittstown, NJ | June | 132.8 | 70.7 | 20.3 | 20.5 | 25.7 | 15.5 | 254.2 |
| July | 175.5 | 76.0 | 24.4 | 24.1 | 29.6 | 19.3 | 267.6 | |
| Aug | 99.3 | 77.4 | 23.3 | 22.1 | 27.6 | 17.7 | 230.0 | |
| Sep | 26.2 | 74.5 | 19.6 | 19.2 | 25.1 | 14.1 | 185.3 | |
| Oct | 137.4 | 77.3 | 14.9 | 13.6 | 18.4 | 9.1 | 110.4 | |
| Nov | 52.6 | 66.6 | 7.8 | 4.2 | 8.9 | −0.5 | 95.7 | |
| Upper Deerfield, NJ | June | 118.4 | 72.6 | 23.0 | 22.9 | 28.6 | 17.3 | 266.0 |
| July | 119.9 | 78.0 | 26.8 | 26.0 | 31.9 | 20.6 | 269.0 | |
| Aug | 54.9 | 79.5 | 25.4 | 24.0 | 30.2 | 19.0 | 217.9 | |
| Sep | 10.2 | 74.2 | 22.5 | 21.3 | 28.6 | 14.7 | 193.0 | |
| Oct | 162.8 | 77.2 | 17.4 | 15.6 | 20.9 | 10.3 | 113.1 | |
| Nov | 32.0 | 69.4 | 9.5 | 5.7 | 11.7 | 0.1 | 98.2 | |
| Weeks after Transplanting | 3 | 6 | 9 | 11 | 13 | 15 | |
|---|---|---|---|---|---|---|---|
| Date | Pittstown | 15 July | 6 August | 28 August | 11 September | 25 September | 9 October |
| Upper Deerfield | 30 July | 19 August | 9 September | 23 September | 7 October | 21 October | |
| Developmental-stage seed | Pittstown | Vegetative | Floral bud | Partial flowering | Full Flowering | Fruit set | Fruit Senescence |
| Upper Deerfield | Vegetative | Vegetative | Floral bud | Partial flowering | Full flowering | Full flowering | |
| Developmental-stage cutting | Pittstown | Vegetative | Partial flowering | Full flowering | Fruit set | Fruit Senescence | Fruit Senescence |
| Upper Deerfield | Vegetative | Vegetative | Floral bud | Partial flowering | Full flowering | Fruit set | |
4. Materials and Methods
4.1. Genotype and Plant Propagation
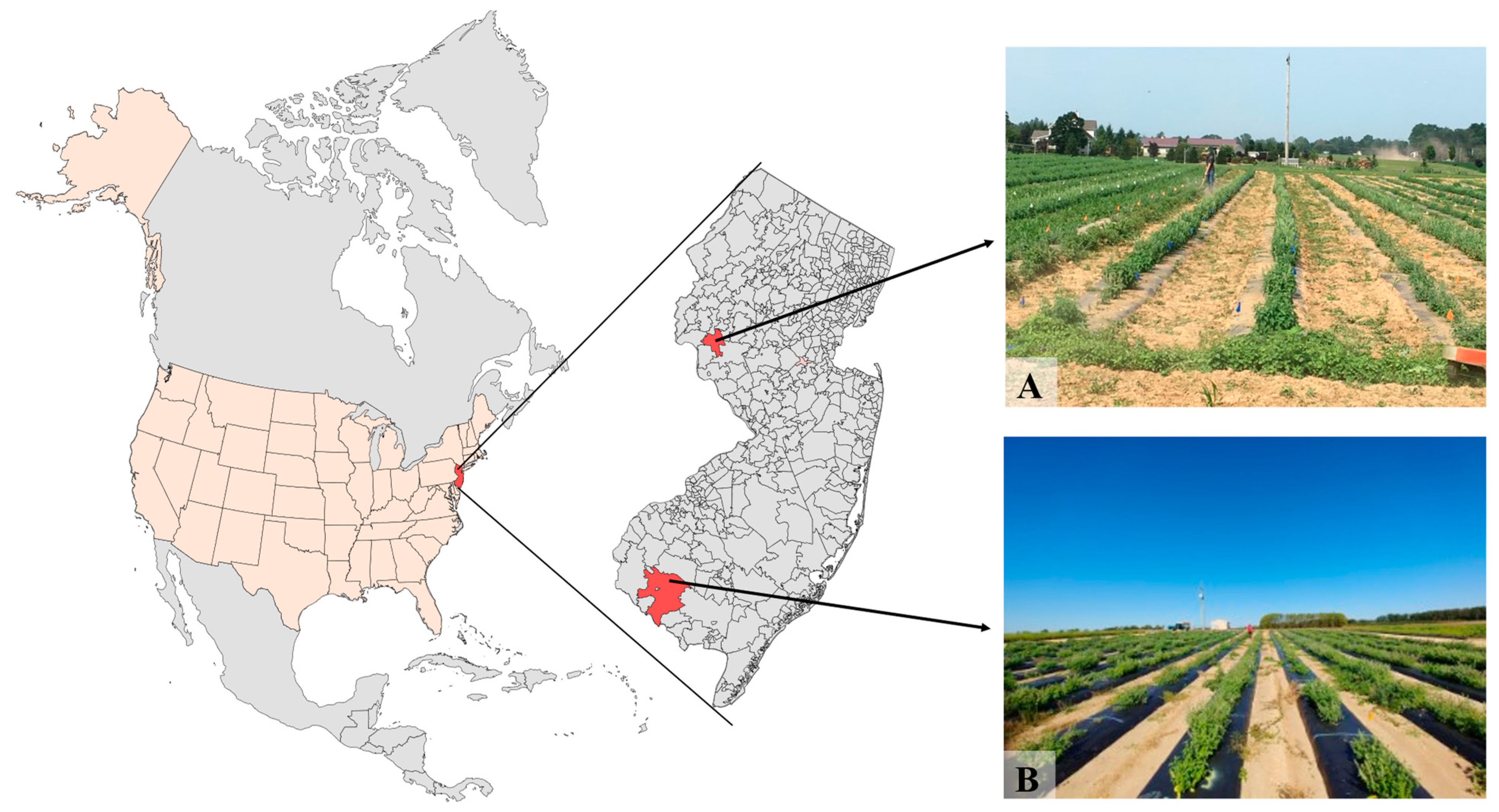
4.2. Field Conditions, Transplanting and Experimental Design
4.3. Determination of Developmental Stages, Harvesting and Postharvest Handling
4.4. Phytochemical and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frezza, C.; Venditti, A.; Serafini, M.; Bianco, A. Phytochemistry, chemotaxonomy, ethnopharmacology, and nutraceutics of Lamiaceae. Stud. Nat. Prod. Chem. 2019, 62, 125–178. [Google Scholar]
- Napoli, E.; Siracusa, L.; Ruberto, G. New tricks for old guys: Recent developments in the chemistry, biochemistry, applications and exploitation of selected species from the Lamiaceae Family. Chem. Biodivers. 2020, 17, e1900677. [Google Scholar] [CrossRef]
- Gomes, E.N.; Allen, K.; Jaworski, K.; Zorde, M.; Lockhart, A.; Besancon, T.; Brown, T.; Reichert, W.; Wu, Q.; Simon, J.E. Catnip (Nepeta cataria L.): Recent Advances in Botany, Horticulture and Production. Medicinal and Aromatic Plants of North America. In Medicinal and Aromatic Plants of North America; Máthé, Á., Ed.; Springer: Cham, Switzerland, 2020; Volume 6, pp. 247–284. [Google Scholar]
- Lichman, B.R.; Godden, G.T.; Hamilton, J.P.; Palmer, L.; Kamileen, M.O.; Zhao, D.; Vaillancourt, B.; Wood, J.C.; Sun, M.; Kinser, T.J.; et al. The evolutionary origins of the cat attractant nepetalactone in catnip. Sci. Adv. 2020, 6, eaba0721. [Google Scholar] [CrossRef]
- Uenoyama, R.; Miyazaki, T.; Adachi, M.; Nishikawa, T.; Hurst, J.L.; Miyazaki, M. Domestic cat damage to plant leaves containing iridoids enhances chemical repellency to pests. iScience 2022, 25, 104455. [Google Scholar] [CrossRef]
- Reichert, W.; Ejercito, J.; Guda, T.; Dong, X.; Wu, Q.L.; Ray, A.; Simon, J.E. Repellency assessment of Nepeta cataria essential oils and isolated nepetalactones on Aedes aegypti. Sci. Rep. 2019, 9, 1524. [Google Scholar] [CrossRef]
- Shi, X.; Wang, C.; Simon, J.E.; Reichert, W.; Wu, Q.L. Repellency of novel catnip oils against the bed bug (Hemiptera: Cimicidae). J. Med. Entomol. 2020, 58, 528–534. [Google Scholar] [CrossRef]
- González, J.; Lockhart, A.; Wu, Q.; Simon, J.E.; Toledo, A. Repellency of novel catnip (Nepeta cataria) cultivar extracts against Ixodes scapularis and Haemaphysalis longicornis (Acari: Ixodida: Ixodidae). Ticks Tick. Borne Dis. 2022, 13, 102046. [Google Scholar] [CrossRef]
- Sharma, A.; Cooper, R.; Bhardwaj, G.; Cannoo, D.S. The genus Nepeta: Traditional uses, phytochemicals and pharmacological properties. J. Ethnopharmacol. 2021, 268, 113679. [Google Scholar] [CrossRef]
- Patel, H.; Gomes, E.N.; Yuan, B.; Lyu, W.; Wu, Q.; Simon, J.E. Investigation of Volatile Iridoid Terpenes in Nepeta cataria L. (Catnip) Genotypes. Molecules 2022, 27, 7057. [Google Scholar] [CrossRef] [PubMed]
- Feaster, J.E.; Scialdone, M.A.; Todd, R.G.; Gonzalez, Y.I.; Foster, J.P.; Hallahan, D.L. Dihydronepetalactones deter feeding activity by mosquitoes, stable flies, and deer ticks. J. Med. Entomol. 2009, 46, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Scialdone, M.A.; Liauw, A.Y. Acetals of Nepetalic Acid and Method of Preparation. U.S. Patent 7,776,912, 17 August 2010. [Google Scholar]
- Bohbot, J.D.; Fu, L.; Le, T.C.; Chauhan, K.R.; Cantrell, C.L.; Dickens, J.C. Multiple activities of insect repellents on odorant receptors in mosquitoes. Med. Vet. Entomol. 2011, 25, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Reichert, W.; Villani, T.; Pan, M.H.; Ho, C.T.; Simon, J.E.; Wu, Q. Phytochemical analysis and anti-inflammatory activity of Nepeta cataria accessions. J. Med. Act. Plants 2018, 7, 19–27. [Google Scholar]
- Gomes, E.N.; Patel, H.; Yuan, B.; Lyu, W.; Juliani, H.R.; Wu, Q.; Simon, J.E. Successive harvests affect the aromatic and polyphenol profiles of novel catnip (Nepeta cataria L.) cultivars in a genotype-dependent manner. Front. Plant Sci. 2023, 14, 329. [Google Scholar] [CrossRef] [PubMed]
- Cipollini, D.; Stevenson, R.; Enright, S.; Eyles, A.; Bonello, P. Phenolic metabolites in leaves of the invasive shrub, Lonicera maackii, and their potential phytotoxic and anti-herbivore effects. J. Chem. Ecol. 2008, 34, 144–152. [Google Scholar] [CrossRef] [PubMed]
- War, A.R.; Paulraj, M.G.; Hussain, B.; Buhroo, A.A.; Ignacimuthu, S.; Sharma, H.C. Effect of plant secondary metabolites on legume pod borer, Helicoverpa armigera. J. Pest. Sci. 2013, 86, 399–408. [Google Scholar] [CrossRef]
- Simmonds, M.S.; Stevenson, P.C.; Hanson, F.E. Rosmarinic acid in Canna generalis activates the medial deterrent chemosensory neurone and deters feeding in the tobacco hornworm Manduca sexta. Physiol. Entomol. 2019, 44, 140–147. [Google Scholar] [CrossRef]
- Reichert, W.; Park, H.C.; Juliani, H.R.; Simon, J.E. ‘CR9’: A New Highly Aromatic Catnip Nepeta cataria L. Cultivar Rich in Z,E-Nepetalactone. HortScience 2016, 51, 588–591. [Google Scholar] [CrossRef]
- Gomes, E.N.; Reichert, W.; Vasilatis, A.A.; Allen, K.A.; Wu, Q.; Simon, J.E. Essential Oil Yield and Aromatic Profile of Lemon Catnip and Lemon-Scented Catnip Selections at Different Harvesting Times. J. Med. Act. Plants 2020, 9, 21–33. [Google Scholar]
- Srivastava, A.; Gupta, S.; Singh, S.; Verma, R.S.; Srivastava, R.K.; Gupta, A.K.; Lal, R.K. Genetic variability and elite line selection for high essential oil and nepetalactone content in catmint (Nepeta cataria L.). Am. J. Plant Sci. 2021, 12, 1135–1154. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Martel, A.B.; Strugnell, C.A. Environmental Factors Regulate Plant Secondary Metabolites. Plants 2023, 12, 447. [Google Scholar] [CrossRef]
- Duda, S.C.; Marghita¸s, L.A.; Dezmirean, D.S.; Duda, M.; Margaoan, R.; Bobis, O. Changes in major bioactive compounds with antioxidant activity of Agastache foeniculum, Lavandula angustifolia, Melissa officinalis and Nepeta cataria: Effect of harvest time and plant species. Ind. Crops Prod. 2015, 77, 499–507. [Google Scholar] [CrossRef]
- Amaral-Baroli, A.; Lago, J.H.G.; de Almeida, C.V.; Almeida, M.; Scotti, M.T.; Leone, G.F.; Soares, M.G.; Cavalari, A.A.; Sartorelli, P. Variability in essential oil composition produced by micropropagated (in vitro), acclimated (ex vitro) and in-field plants of Ocimum basilicum (Lamiaceae). Ind. Crops Prod. 2016, 86, 180–185. [Google Scholar] [CrossRef]
- Said-Al Ahl, H.A.; Sabra, A.S.; Hegazy, M.H. Salicylic acid improves growth and essential oil accumulation in two Nepeta cataria chemotypes under water stress conditions. Ital. J. Agrometeorol. 2016, 21, 25–36. [Google Scholar]
- Gomes, E.N.; Moterle, D.; Biasi, L.A.; Koehler, H.S.; Kanis, L.A.; Deschamps, C. Plant densities and harvesting times on productive and physiological aspects of Stevia rebaudiana Bertoni grown in southern Brazil. An. Acad. Bras. Cien. 2018, 90, 3249–3264. [Google Scholar] [CrossRef] [PubMed]
- Mirjalili, M.H.; Salehi, P.; Vala, M.M.; Ghorbanpour, M. The effect of drying methods on yield and chemical constituents of the essential oil in Lavandula angustifolia Mill. (Lamiaceae). Plant Physiol. Rep. 2019, 24, 96–103. [Google Scholar] [CrossRef]
- Schultz, G.; Simbro, E.; Belden, J.; Zhu, J.; Coats, J. Catnip, Nepeta cataria (Lamiales: Lamiaceae)—A closer look: Seasonal occurrence of nepetalactone isomers and comparative repellency of three terpenoids to insects. Environ. Entomol. 2004, 33, 1562–1569. [Google Scholar] [CrossRef]
- Bourrel, C.; Perineau, F.; Michel, G.; Bessiere, J.M. Catnip (Nepeta cataria L.) essential oil: Analysis of chemical constituents, bacteriostatic and fungistatic properties. J. Essent. Oil Res. 1993, 5, 159–167. [Google Scholar] [CrossRef]
- Mohammadi, S.; Saharkhiz, M.J. Changes in essential oil content and composition of catnip (Nepeta cataria L.) during different developmental stages. J. Essent. Oil-Bear. Plants 2011, 14, 396–400. [Google Scholar] [CrossRef]
- Ibrahim, M.E.; El-Sawi, S.A.; Ibrahim, F.M. Nepeta cataria L., one of the promising aromatic plants in Egypt: Seed germination, growth and essential oil production. J. Mater. Environ. Sci. 2017, 8, 1990–1995. [Google Scholar]
- Hilaire, R.S. Propagation of catnip by terminal and single-node cuttings. J. Environ. Hort. 2003, 21, 20–23. [Google Scholar] [CrossRef]
- Hilaire, R.S.; Hockman, A.W.; Chavez, S.M. Propagation and irrigation regime affect the development of catnip. Acta Hortic. 2004, 629, 321–327. [Google Scholar] [CrossRef]
- Allen, K.A.; Nunes Gomes, E.; Patel, H.K.; Simon, J.E. Vegetative propagation of Nepeta cataria and the inhibitory effects of its essential oil on the adventitious rooting of cultivated lamiaceae species. J. Med. Act. Plants 2023, 12, 18–31. [Google Scholar]
- Falk, C.L.; Voorthuizen, H.V.; Wall, M.M.; Guldan, S.J.; Martin, C.A.; Kleitz, K.M. An economic analysis of transplanting versus direct seeding of selected medicinal herbs in New Mexico. J. Herbs Spices Med. Plants 2000, 7, 15–29. [Google Scholar] [CrossRef]
- McKey, D.; Elias, M.; Pujol, B.; Duputié, A. The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 2010, 186, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Heybroek, H.M.; Visser, T. Juvenility in fruit growing and forestry. Acta Hortic. 1976, 56, 71–80. [Google Scholar] [CrossRef]
- Albrecht, U.; Bordas, M.; Lamb, B.; Meyering, B.; Bowman, K.D. Influence of propagation method on root architecture and other traits of young citrus rootstock plants. HortScience 2017, 52, 1569–1576. [Google Scholar] [CrossRef]
- Heide, O.M. Juvenility, maturation and rejuvenation in plants: Adventitious bud formation as a novel rejuvenation process. J. Hortic. Sci. Biotechnol. 2019, 94, 2–11. [Google Scholar] [CrossRef]
- Pokhrel, S.; Meyering, B.; Bowman, K.D.; Albrecht, U. Horticultural attributes and root architectures of field-grown ‘Valencia’trees grafted on different rootstocks propagated by seed, cuttings, and tissue culture. HortScience 2021, 56, 163–172. [Google Scholar] [CrossRef]
- Barra, A. Factors affecting chemical variability of essential oils: A review of recent developments. Nat. Prod. Commun. 2009, 4, 1934578X0900400827. [Google Scholar] [CrossRef]
- Liebelt, D.J.; Jordan, J.T.; Doherty, C.J. Only a matter of time: The impact of daily and seasonal rhythms on phytochemicals. Phytochem. Rev. 2019, 18, 1409–1433. [Google Scholar]
- Zhao, Y.; Liu, G.; Yang, F.; Liang, Y.; Gao, Q.; Xiang, C.; Li, X.; Yang, R.; Zhang, G.; Jiang, H.; et al. Multilayered regulation of secondary metabolism in medicinal plants. Mol. Hort. 2023, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Legris, M.; Nieto, C.; Sellaro, R.; Prat, S.; Casal, J.J. Perception and signalling of light and temperature cues in plants. Plant J. 2017, 90, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Perrella, G.; Vellutini, E.; Zioutopoulou, A.; Patitaki, E.; Headland, L.R.; Kaiserli, E. Let it bloom: Cross-talk between light and flowering signaling in Arabidopsis. Physiol. Plant. 2020, 169, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Lee, J.H.; Kim, W.; Jung, H.S.; Huijser, P.; Ahn, J.H. The microRNA156-SQUAMOSA Promoter Binding Protein-LIKE3 module regulates ambient temperature-responsive flowering via Flowering Locus T in Arabidopsis. Plant Physiol. 2012, 159, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, C.A.; Anderssen, R.S.; Robertson, M.; Finnegan, E.J. How is FLC repression initiated by cold? Trends Plant Sci. 2015, 20, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Deschamps, C.; Gang, D.; Dudareva, N.; Simon, J.E. Developmental regulation of phenylpropanoid biosynthesis in leaves and glandular trichomes of basil (Ocimum basilicum L.). Int. J. Plant Sci. 2006, 67, 447–454. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R. Phenolic Compounds from New Natural Sources—Plant Genotype and Ontogenetic Variation. Molecules 2023, 28, 1731. [Google Scholar] [CrossRef] [PubMed]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef]
- Hornok, L.; Domokos, J.; Hethelyi, E. Effect of harvesting time on the production of Nepeta cataria var. citriodora Balb. Acta Hortic. 1992, 306, 290–294. [Google Scholar] [CrossRef]
- Marino, S.; Ahmad, U.; Ferreira, M.I.; Alvino, A. Evaluation of the effect of irrigation on biometric growth, physiological response, and essential oil of Mentha spicata (L.). Water 2019, 11, 2264. [Google Scholar] [CrossRef]
- Copolovici, L.; Copolovici, D.M.; Moisa, C.; Lupitu, A. Antagonist Temperature Variation Affects the Photosynthetic Parameters and Secondary Metabolites of Ocimum basilicum L. and Salvia officinalis L. Plants 2022, 11, 1806. [Google Scholar] [CrossRef]
- Manukyan, A. Effect of growing factors on productivity and quality of lemon catmint, lemon balm and sage under soilless greenhouse production: I. Drought stress. Med. Arom. Plant Sci. Biotechnol. 2011, 5, 119–125. [Google Scholar]
- Chalchat, J.C.; Lamy, J. Chemical composition of the essential oil isolated from wild catnip Nepeta cataria L. cv. Citriodora from the Drôme region of France. J. Essent. Oil Res. 1997, 9, 527–532. [Google Scholar] [CrossRef]
- Bondarev, N.I.; Sukhanova, M.A.; Reshetnyak, O.V.; Nosov, A.M. Steviol glycoside content in different organs of Stevia rebaudiana and its dynamics during ontogeny. Biol. Plant. 2003, 47, 261–264. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, S.; Han, Y.; Yuan, H.; Gu, C.; Wang, Z. Environmental cues induce changes of steviol glycosides contents and transcription of corresponding biosynthetic genes in Stevia rebaudiana. Plant Physiol. Biochem. 2015, 86, 174–180. [Google Scholar] [CrossRef] [PubMed]
- McConkey, M.E.; Gershenzon, J.; Croteau, R.B. Developmental regulation of monoterpene biosynthesis in the glandular trichomes of peppermint. Plant Physiol. 2000, 122, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Renu, I.K.; Haque, I.; Kumar, M.; Poddar, R.; Bandopadhyay, R.; Rai, A.; Mukhopadhyay, K. Characterization and functional analysis of eugenol O-methyltransferase gene reveal metabolite shifts, chemotype specific differential expression and developmental regulation in Ocimum tenuiflorum L. Mol. Biol. Rep. 2014, 41, 1857–1870. [Google Scholar] [CrossRef]
- Aničić, N.; Matekalo, D.; Skorić, M.; Pećinar, I.; Brkušanin, M.; Živković, J.N.; Mišić, D. Trichome-specific and developmentally regulated biosynthesis of nepetalactones in leaves of cultivated Nepeta rtanjensis plants. Ind. Crops Prod. 2018, 117, 347–358. [Google Scholar] [CrossRef]
- Simon, J.E.; Reichert, W.; Wu, Q. Catnip Cultivar ‘CR3’. U.S. Patent 10,512,231, 24 December 2019. [Google Scholar]
- Fassio, C.; Cautin, R.; Pérez-Donoso, A.; Bonomelli, C.; Castro, M. Propagation techniques and grafting modify the morphological traits of roots and biomass allocation in avocado trees. HortTechnology 2016, 26, 63–69. [Google Scholar] [CrossRef]
- Carr, M.K.V. Responses of seedling tea bushes and their clones to water stress. Exp. Agric. 1977, 13, 317–324. [Google Scholar] [CrossRef]
- Blake, T.J.; Filho, W.S. Drought tolerance, growth partitioning and vigor in eucalypt seedlings and rooted cuttings. Tree Physiol. 1988, 4, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Plant Propagation: Principles and Practices; Prentice-Hall Inc.: Upper Saddle River, NJ, USA, 1997. [Google Scholar]
- Hadi, N.; Sefidkon, F.; Shojaeiyan, A.; Šiler, B.; Jafari, A.A.; Aničić, N.; Mišić, D. Phenolics’ composition in four endemic Nepeta species from Iran cultivated under experimental field conditions: The possibility of the exploitation of Nepeta germplasm. Ind. Crops Prod. 2017, 95, 475–484. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, Q.; Liu, L.; Liao, L.; Zhu, Z. Influence of fertilization and drought stress on the growth and production of secondary metabolites in Prunella vulgaris L. J. Med. Plants Res. 2011, 5, 1749–1755. [Google Scholar]
- Zafar-ul-Hye, M.; Akbar, M.N.; Iftikhar, Y.; Abbas, M.; Zahid, A.; Fahad, S.; Danish, S. Rhizobacteria inoculation and caffeic acid alleviated drought stress in lentil plants. Sustainability 2021, 13, 9603. [Google Scholar] [CrossRef]
- Riaz, U.; Kharal, M.A.; Murtaza, G.; Zaman, Q.; Javaid, S.; Malik, H.A.; Abbas, Z. Prospective roles and mechanisms of caffeic acid in counter plant stress: A mini review. Pak. J. Agric. Res. 2019, 32, 8. [Google Scholar] [CrossRef]
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible flavone in oats (Avena sativa) is a novel defense against plant-parasitic nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef]
- Righini, S.; Rodriguez, E.J.; Berosich, C.; Grotewold, E.; Casati, P.; Falcone Ferreyra, M.L. Apigenin produced by maize flavone synthase I and II protects plants against UV-B-induced damage. Plant Cell Environ. 2019, 42, 495–508. [Google Scholar] [CrossRef]
- Mechri, B.; Tekaya, M.; Hammami, M.; Chehab, H. Effects of drought stress on phenolic accumulation in greenhouse-grown olive trees (Olea europaea). Biochem. Syst. Ecol. 2020, 92, 104112. [Google Scholar] [CrossRef]
- Yun, D.Y.; Kang, Y.G.; Kim, M.; Kim, D.; Kim, E.H.; Hong, Y.S. Metabolomic understanding of pod removal effect in soybean plants and potential association with their health benefit. Food Res. Int. 2020, 138, 109797. [Google Scholar] [CrossRef]
- Xue, J.; Lu, D.; Wang, S.; Lu, Z.; Liu, W.; Wang, X.; He, X. Integrated transcriptomic and metabolomic analysis provides insight into the regulation of leaf senescence in rice. Sci. Rep. 2021, 11, 14083. [Google Scholar] [CrossRef]
- Goławska, S.; Łukasik, I.; Goławski, A.; Kapusta, I.; Janda, B. Alfalfa (Medicago sativa L.) apigenin glycosides and their effect on the pea aphid (Acyrthosiphon pisum). Pol. J. Environ. Stud. 2010, 19, 913–919. [Google Scholar]
- Rutgers NJ Weather Network. Available online: https://www.njweather.org/ (accessed on 24 December 2019).
- Silva, F.D.A.S.; Azevedo, C.A.V. The Assistat Software Version 7.7 and its use in the analysis of experimental data. Afr. J. Agric. Res. 2016, 11, 3733–3740. [Google Scholar]







| Compounds | RT * (min) | Precursor (m/z) | Frag | Quant (m/z) | CE (eV) | Qual 1 (m/z) | CE (eV) | Ratio | Qual 2 (m/z) | CE (eV) | Ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Z,E-NL | 3.57 | 167.0 | 95 | 55.1 | 34 | 77.0 | 38 | 0.99 | 43.1 | 30 | 0.87 |
| E,Z-NL | 3.89 | 167.0 | 70 | 77.0 | 41 | 43.1 | 30 | 0.78 | 81.1 | 13 | 0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, E.N.; Yuan, B.; Patel, H.K.; Lockhart, A.; Wyenandt, C.A.; Wu, Q.; Simon, J.E. Implications of the Propagation Method for the Phytochemistry of Nepeta cataria L. throughout a Growing Season. Molecules 2024, 29, 2001. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29092001
Gomes EN, Yuan B, Patel HK, Lockhart A, Wyenandt CA, Wu Q, Simon JE. Implications of the Propagation Method for the Phytochemistry of Nepeta cataria L. throughout a Growing Season. Molecules. 2024; 29(9):2001. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29092001
Chicago/Turabian StyleGomes, Erik Nunes, Bo Yuan, Harna K. Patel, Anthony Lockhart, Christian A. Wyenandt, Qingli Wu, and James E. Simon. 2024. "Implications of the Propagation Method for the Phytochemistry of Nepeta cataria L. throughout a Growing Season" Molecules 29, no. 9: 2001. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29092001