

Enrichment and Evaluation of Antitumor Properties of Total Flavonoids from Juglans mandshurica Maxim

Abstract
:1. Introduction
2. Results and Discussion
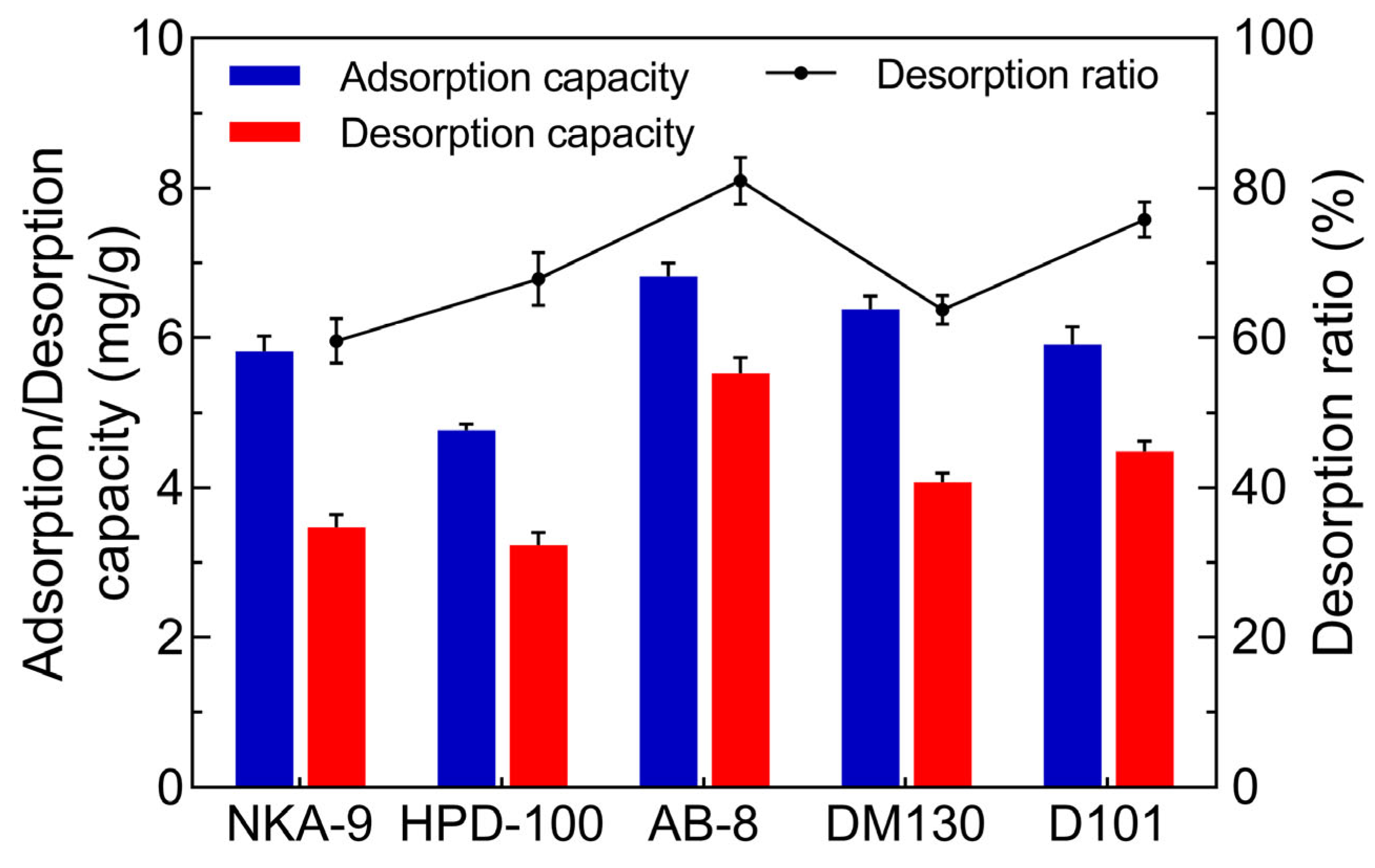
2.1. Screening the Suitable Macroporous Resin for JMFs Purification
2.2. Adsorption Kinetics of the AB-8 Resin
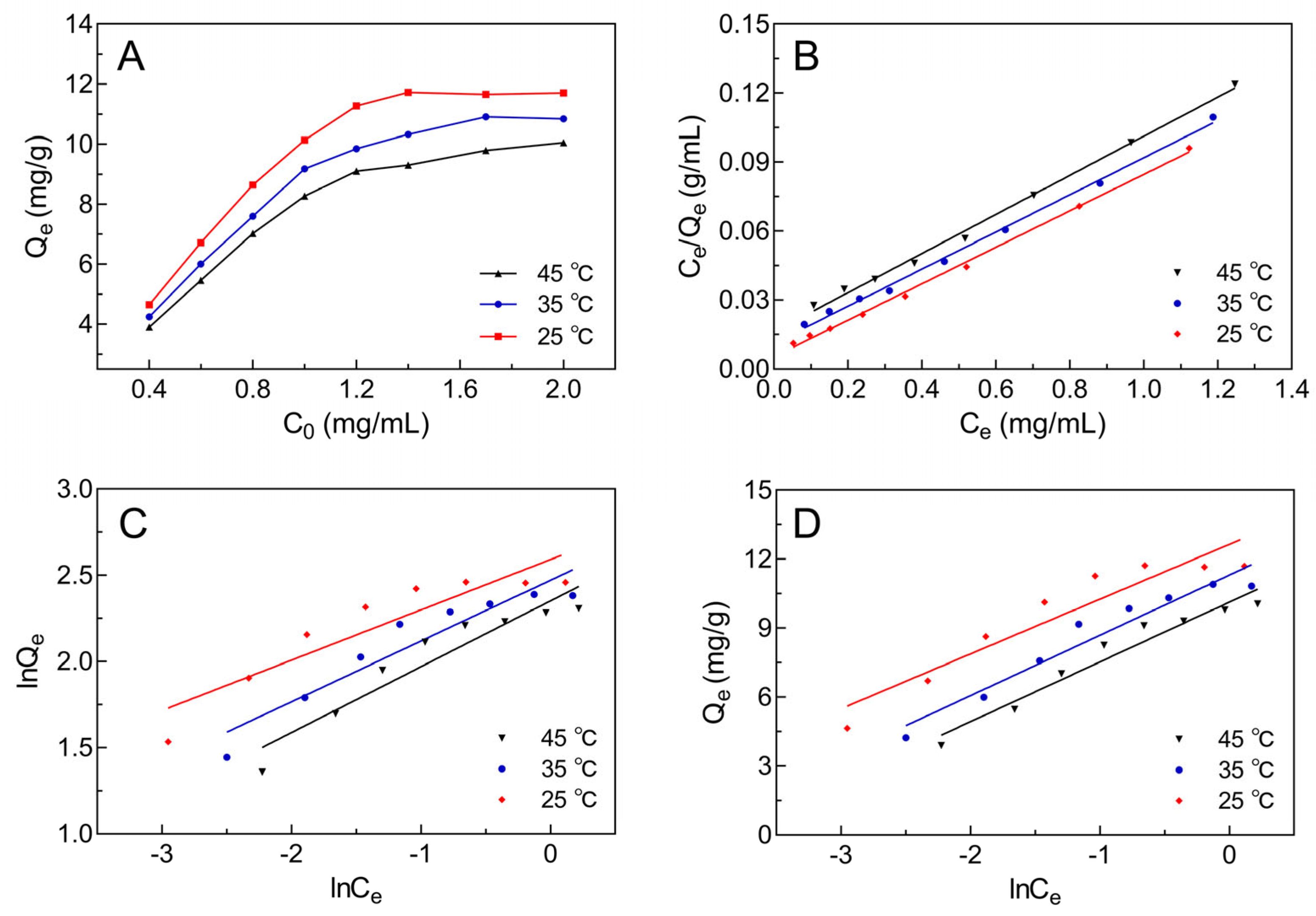
2.3. Adsorption Isotherm Curve of Selected Macroporous Resin
2.4. Dynamic Adsorption and Desorption Curves on the AB-8 Resin towards JMFs
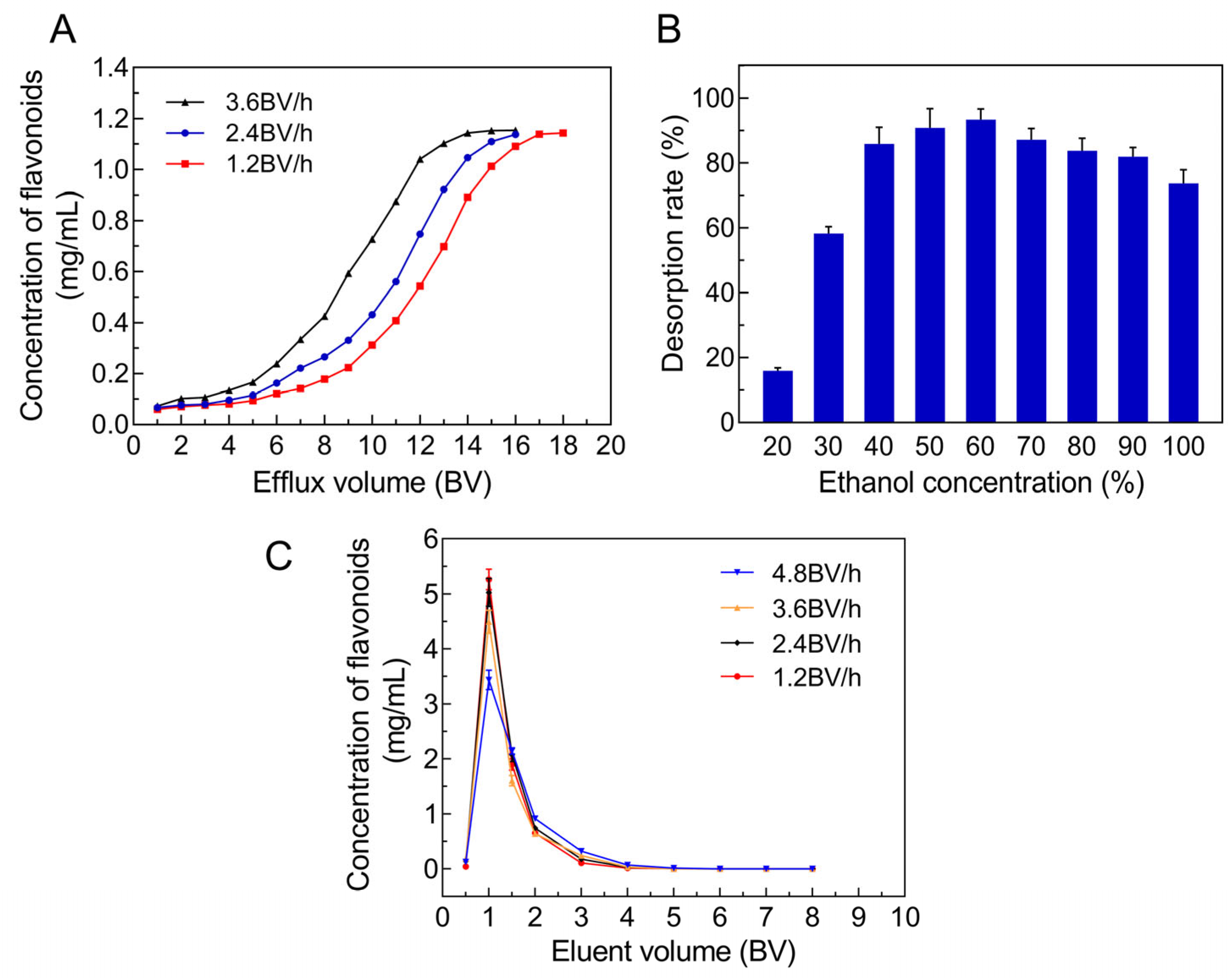
2.4.1. Screening the Suitable Adsorption Conditions
2.4.2. Screening the Suitable Desorption Conditions
2.4.3. Validation Experiments
2.5. FTIR Analysis of Enriched JMFs
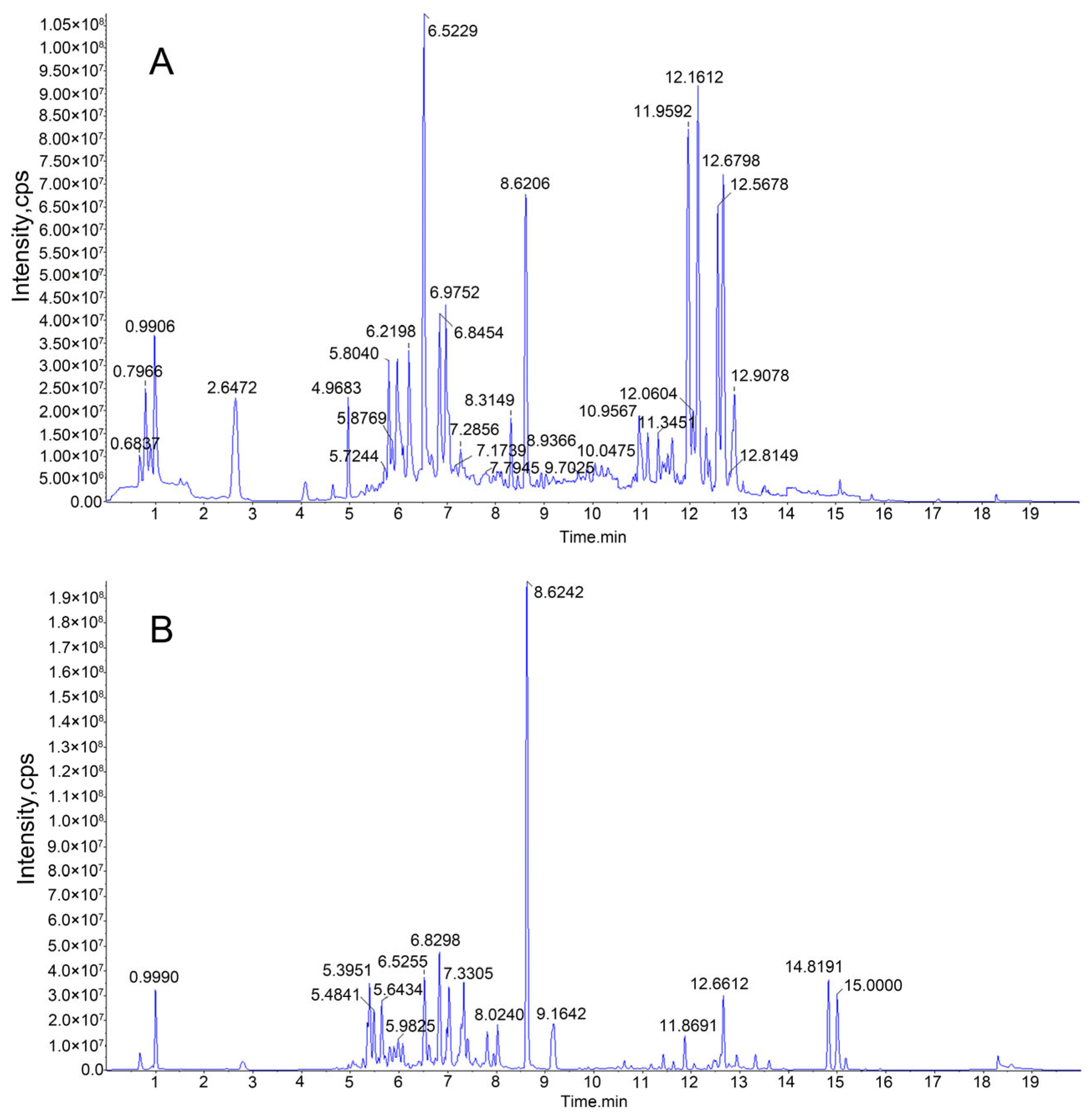
2.6. HPLC-MS/MS Analysis of Enriched JMFs
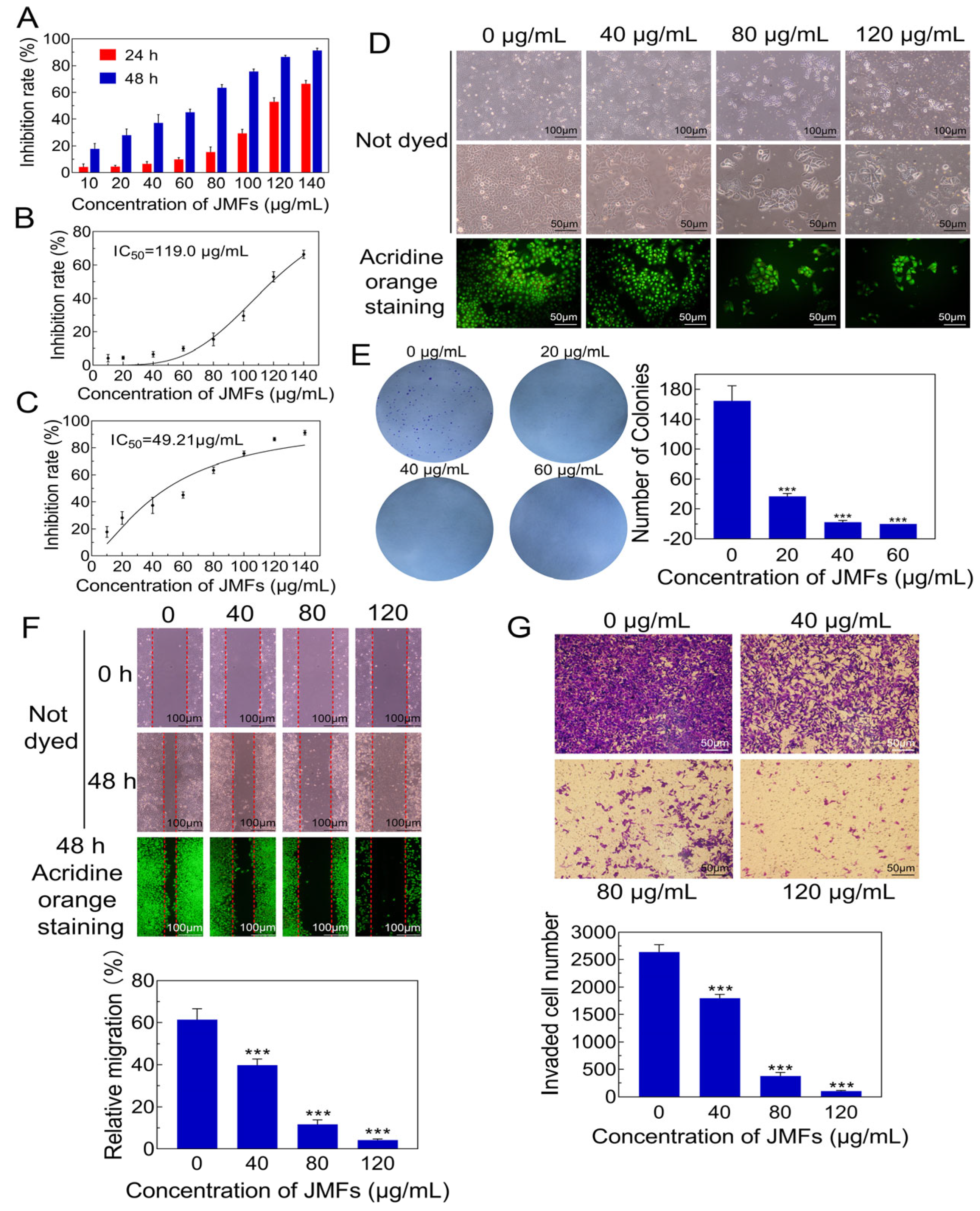
2.7. Anticancer Potential of Purified JMFs
2.7.1. Anti-Proliferation Effect of JMFs on HepG2 Cells
2.7.2. Morphological Changes in JMFs-Exposed HepG2 Cells
2.7.3. Inhibitory Potential of JMFs on Colony Formation of HepG2 Cells
2.7.4. JMFs Inhibited Migration and Invasion Abilities of HepG2 Cells
2.7.5. JMFs Exposure Triggered S Phase Arrest in HepG2 Cells
2.7.6. Changes in ROS Levels in HepG2 Cells Treated with JMFs
2.7.7. Induction of Apoptosis of HepG2 Cells by JMFs
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Preparation of the Crude Flavonoid Extracts from J. mandshurica
3.3. Static Adsorption and Desorption Tests
3.4. Adsorption Kinetics
3.5. Adsorption Isotherms
3.6. Dynamic Adsorption and Desorption Tests
3.7. Fourier Transform Infrared Spectroscopy Analyses of JMFs
3.8. High-Performance Liquid Chromatography–Mass Spectrometry (HPLC-MS/MS)
3.9. Antitumor Efficiency of JMFs
3.9.1. Culture Conditions of Cells Used in the Present Study
3.9.2. Cell Viability Detection by CCK8 Kit
3.9.3. Observation of Cell Morphology
3.9.4. Colony-Forming Assay
3.9.5. Migration and Invasion Assay
3.9.6. Cell Cycle Analysis
3.9.7. Determination of Reactive Oxygen Species (ROS)
3.9.8. Apoptosis Was Detected by Hoechst 33342 Staining
3.9.9. Annexin V-FITC/PI Staining
3.10. Western Blot
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Polkowska-Pruszyńska, B.; Rawicz-Pruszyński, K.; Ciseł, B.; Sitarz, R.; Polkowska, G.; Krupski, W.; Polkowski, W.P. Liver metastases from gastric carcinoma: A Case report and review of the literature. Curr. Probl. Cancer 2017, 41, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Reddy, N.D.; Shoja, M.H.; Biswas, S.; Nayak, P.G.; Kumar, N.; Rao, C.M. An appraisal of cinnamyl sulfonamide hydroxamate derivatives (HDAC inhibitors) for anti-cancer, anti-angiogenic and anti-metastatic activities in human cancer cells. Chem. Biol. Interact. 2016, 253, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tao, L.; Ni, T.; Gu, H.; Jin, F.; Dai, X.; Feng, J.; Ding, Y.; Xiao, W.; Guo, S.; et al. Anticancer efficacy of the ethyl acetate extract from the traditional Chinese medicine herb Celastrus orbiculatus against human gastric cancer. J. Ethnopharmacol. 2017, 205, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric cancer: Descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ji, F. Downregulation of microRNA-17-5p inhibits drug resistance of gastric cancer cells partially through targeting p21. Oncol. Lett. 2018, 15, 4585–4591. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Li, H.; Sun, D.; Chen, W. Cancer burden of major cancers in China: A need for sustainable actions. Cancer Commun. 2020, 40, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, H. Precision diagnosis and treatment of liver cancer in China. Cancer Lett. 2018, 412, 283–288. [Google Scholar] [CrossRef]
- Shafabakhsh, R.; Yousefi, B.; Asemi, Z.; Nikfar, B.; Mansournia, M.A.; Hallajzadeh, J. Chitosan: A compound for drug delivery system in gastric cancer-a review. Carbohydr. Polym. 2020, 242, 116403. [Google Scholar] [CrossRef]
- Abbasi, B.A.; Iqbal, J.; Ahmad, R.; Bibi, S.; Mahmood, T.; Kanwal, S.; Bashir, S.; Gul, F.; Hameed, S. Potential phytochemicals in the prevention and treatment of esophagus cancer: A green therapeutic approach. Pharmacol. Rep. 2019, 71, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Tsouh Fokou, P.V.; Yamthe, L.; Tali, B.T.; Adetunji, C.O.; Rahavian, A.; Mudau, F.N.; Martins, N.; Martorell, M.; Rodrigues, C.F.; et al. Insights on the anticancer potential of plant-food bioactives: A key focus to prostate cancer. Cell. Mol. Biol. 2020, 66, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.L.; Yu, X.F.; Qu, S.C.; Zhang, R.; Qu, X.R.; Chen, Y.P.; Ma, X.Y.; Sui, D.Y. Anti-proliferative effect of Juglone from Juglans mandshurica Maxim on human leukemia cell HL-60 by inducing apoptosis through the mitochondria-dependent pathway. Eur. J. Pharmacol. 2010, 645, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, W.; Zhang, H.; Liu, J.; Shi, B.; Dai, W.; Liu, K.; Zhang, H. Diversity in Fruit Morphology and Nutritional Composition of Juglans mandshurica Maxim in Northeast China. Front. Plant Sci. 2022, 13, 820457. [Google Scholar] [CrossRef] [PubMed]
- Bayram, D.; Özgöçmen, M.; Armagan, I.; Sevimli, M.; Türel, G.Y.; Şenol, N. Investigation of apoptotic effect of juglone on CCL-228-SW 480 colon cancer cell line. J. Cancer Res. Ther. 2019, 15, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Du, X.; Sun, G.; Meng, Y.; Wang, W. Comparison of the chemical profiles of fresh-raw and dry-processed Juglans mandshurica. J. Sep. Sci. 2017, 40, 646–662. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; Wang, Z.; Yang, Y.; Ji, Y.; Lv, H.; Han, K.; Liu, D.; Shang, X.; He, X.; Zeng, N. Juglans mandshurica Maxim.: A review of its traditional usages, phytochemical constituents, and pharmacological properties. Front. Pharmacol. 2021, 11, 569800. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, J.; Liu, A.; Xiao, B.; Li, S.; Wen, Z.; Lu, Y.; Du, G. Research progress of the antiviral bioactivities of natural flavonoids. Nat. Prod. Bioprospect. 2020, 10, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Jucá, M.M.; Cysne Filho, F.; de Almeida, J.C.; Mesquita, D.; Barriga, J.; Dias, K.; Barbosa, T.M.; Vasconcelos, L.C.; Leal, L.; Ribeiro, J.E.; et al. Flavonoids: Biological activities and therapeutic potential. Nat. Prod. Res. 2020, 34, 692–705. [Google Scholar] [CrossRef]
- Ren, J.; Zheng, Y.; Lin, Z.; Han, X.; Liao, W. Macroporous resin purification and characterization of flavonoids from Platycladus orientalis (L.) Franco and their effects on macrophage inflammatory response. Food Funct. 2017, 8, 86–95. [Google Scholar] [CrossRef]
- Selvakumar, P.; Badgeley, A.; Murphy, P.; Anwar, H.; Sharma, U.; Lawrence, K.; Lakshmikuttyamma, A. Flavonoids and other polyphenols act as epigenetic modifiers in breast cancer. Nutrients 2020, 12, 761. [Google Scholar] [CrossRef] [PubMed]
- Pham, D.C.; Nguyen, H.C.; Nguyen, T.L.; Ho, H.L.; Trinh, T.K.; Riyaphan, J.; Weng, C.F. Optimization of ultrasound-assisted extraction of flavonoids from Celastrus hindsii leaves using response surface methodology and evaluation of their antioxidant and antitumor activities. Biomed. Res. Int. 2020, 3497107. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Raffa, D.; Maggio, B.; Raimondi, M.V.; Plescia, F.; Daidone, G. Recent discoveries of anticancer flavonoids. Eur. J. Med. Chem. 2017, 142, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wang, W.; Zhang, R. Novel natural product therapeutics targeting both inflammation and cancer. Chin. J. Nat. Med. 2017, 15, 401–416. [Google Scholar] [CrossRef]
- Lou, L.L.; Cheng, Z.Y.; Guo, R.; Yao, G.D.; Song, S.J. Alkaloids from Juglans Mandshurica maxim induce distinctive cell death in hepatocellular carcinoma cells. Nat. Prod. Res. 2019, 33, 911–914. [Google Scholar] [CrossRef]
- Cui, J.; Duan, X.; Ke, L.; Pan, X.; Liu, J.; Song, X.; Ma, W.; Zhang, W.; Liu, Y.; Fan, Y. Extraction, purification, structural character and biological properties of propolis flavonoids: A review. Fitoterapia 2022, 157, 105106. [Google Scholar] [CrossRef]
- Chen, X.; Wang, H.; Huang, X.; Xia, S.; Chen, C.; Nie, Q.; Nie, S. Efficient enrichment of total flavonoids from kale (Brassica oleracea L. var acephala L.) extracts by NKA-9 resin and antioxidant activities of flavonoids extract in vitro. Food Chem. 2022, 374, 131508. [Google Scholar] [CrossRef]
- Hou, M.; Hu, W.; Xiu, Z.; Shi, Y.; Hao, K.; Cao, D.; Guan, Y.; Yin, H. Efficient enrichment of total flavonoids from Pteris ensiformis Burm. extracts by macroporous adsorption resins and in vitro evaluation of antioxidant and antiproliferative activities. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2020, 1138, 121960. [Google Scholar] [CrossRef]
- Noroozi, B.; Sorial, G.A.; Bahrami, H.; Arami, M. Equilibrium and kinetic adsorption study of a cationic dye by a natural adsorbent--silkworm pupa. J. Hazard. Mater. 2007, 139, 167–174. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.; Zhao, T.; Li, F.; Zhang, M.; Li, J.; Zou, Y.; Wang, W.; Cobbina, S.J.; Wu, X.; et al. Adsorption properties of macroporous adsorbent resins for separation of anthocyanins from mulberry. Food Chem. 2016, 194, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Hu, W.; Xiu, Z.; Jiang, A.; Men, L.; Hao, K.; Sun, X.; Cao, D. Preparative purification of total flavonoids from Sophora tonkinensis gagnep. by macroporous resin column chromatography and comparative analysis of flavonoid profiles by HPLC-PAD. Molecules 2019, 24, 3200. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Labreche, F.; Wu, C.; Fan, G.; Li, T.; Dou, J.; Zhu, J. Ultrasound-assisted adsorption/desorption of jujube peel flavonoids using macroporous resins. Food Chem. 2022, 368, 130800. [Google Scholar] [CrossRef]
- Li, Q.L.; Xin, W.X.; Zhong, L.K.; Fang, L.; Cao, G.; Huang, P. A study on the anti-tumor mechanism of total flavonoids from Radix Tetrastigmae against additional cell line based on COX-2-mediated Wnt/β-catenin signaling pathway. Oncotarget 2017, 8, 54304–54319. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.; Jiang, L.J.; Zhen, H.S.; Huang, F.Y.; Zhen, D.D.; Ye, M.F.; Meng, X.Y.; Liu, Y.Y.; Qin, X.J. Promotion of HepG2 cell apoptosis by Sedum emarginatum Migo and the mechanism of action. BMC Complement. Med. Ther. 2022, 22, 31. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Deng, G.; Li, Y.; Huang, H.; Sun, X.; Shi, H.; Yao, X.; Gao, L.; Ju, Y.; Luo, M. Actinidia chinensis Planch prevents proliferation and migration of gastric cancer associated with apoptosis, ferroptosis activation and mesenchymal phenotype suppression. Biomed. Pharmacother. 2020, 126, 110092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.D.; Yu, L.; Wang, P.; Kou, P.; Li, J.; Wang, L.T.; Wang, W.; Yao, L.P.; Zhao, X.H.; Fu, Y.J. Inotodiol inhibits cells migration and invasion and induces apoptosis via p53-dependent pathway in HeLa cells. Phytomedicine 2019, 60, 152957. [Google Scholar] [CrossRef] [PubMed]
- Zaher, A.F.; Abuel-Maaty, S.M.; El-Nassan, H.B.; Amer, S.A.; Abdelghany, T.M. Synthesis, antitumor screening and cell cycle analysis of novel benzothieno[3,2-b]pyran derivatives. J. Enzyme Inhib. Med. Chem. 2016, 31, 145–153. [Google Scholar] [CrossRef]
- Zhong, B.; Shi, D.; Wu, F.; Wang, S.; Hu, H.; Cheng, C.; Qing, X.; Huang, X.; Luo, X.; Zhang, Z.; et al. Dynasore suppresses cell proliferation, migration, and invasion and enhances the antitumor capacity of cisplatin via STAT3 pathway in osteosarcoma. Cell Death Dis. 2019, 10, 687. [Google Scholar] [CrossRef]
- Ji, P.; Huang, H.; Yuan, S.; Wang, L.; Wang, S.; Chen, Y.; Feng, N.; Veroniaina, H.; Wu, Z.; Wu, Z.; et al. ROS-mediated apoptosis and anticancer effect achieved by artesunate and auxiliary Fe(II) released from ferriferous oxide-containing recombinant apoferritin. Adv. Healthc. Mater. 2019, 8, e1900911. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, H.S.; Seo, Y.R. Understanding of ROS-inducing strategy in anticancer therapy. Oxid. Med. Cell. Longev. 2019, 5381692. [Google Scholar] [CrossRef]
- Kang, R.; Li, R.; Dai, P.; Li, Z.; Li, Y.; Li, C. Deoxynivalenol induced apoptosis and inflammation of IPEC-J2 cells by promoting ROS production. Environ. Pollut. 2019, 251, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liao, A.M.; Thakur, K.; Zhang, J.G.; Huang, J.H.; Wei, Z.J. Licochalcone B extracted from Glycyrrhiza uralensis Fisch induces apoptotic effects in human hepatoma cell HepG2. J. Agric. Food Chem. 2019, 67, 3341–3353. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.G. Mechanisms of caspase activation and inhibition during apoptosis. Mol. Cell 2002, 9, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Jin, C.Y.; Kim, G.Y.; Jeong, Y.K.; Kim, W.J.; Choi, Y.H. Induction of apoptosis by ethanol extract of Prunus mume in U937 human leukemia cells through activation of caspases. Oncol. Rep. 2011, 26, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Kogo, J.; Takeba, Y.; Kumai, T.; Kitaoka, Y.; Matsumoto, N.; Ueno, S.; Kobayashi, S. Involvement of TNF-α in glutamate-induced apoptosis in a differentiated neuronal cell line. Brain Res. 2006, 1122, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Bersch, U.; Edelmann, B.; Tchikov, V.; Winoto-Morbach, S.; Schutze, S. Compartmentalization of TNF-receptor 1 signaling: TNF-R1-associated caspase-8 mediates activation of acid sphingomyelinase in late endosomes. Adv. Exp. Med. Biol. 2011, 691, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Seon, M.R.; Lim, S.S.; Choi, H.J.; Park, S.Y.; Cho, H.J.; Kim, J.K.; Kim, J.; Kwon, D.Y.; Park, J.H. Isoangustone A present in hexane/ethanol extract of Glycyrrhiza uralensis induces apoptosis in DU145 human prostate cancer cells via the activation of DR4 and intrinsic apoptosis pathway. Mol. Nutr. Food Res. 2010, 54, 1329–1339. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, J.; Li, Y.J.; Zheng, Y.F.; Li, P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chem. 2013, 141, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Ow, Y.P.; Green, D.R.; Hao, Z.; Mak, T.W. Cytochrome c: Functions beyond respiration. Nat. Rev. Mol. Cell Biol. 2008, 9, 532–542. [Google Scholar] [CrossRef]
- Ge, Y.; Gao, Q.; Yan, F.; Zhang, X.; Liu, Y.; Zhou, Y. Su Yang Decoction induces human colon carcinoma cell apoptosis by activating caspases. Oncol. Lett. 2019, 17, 422–431. [Google Scholar] [CrossRef]
- Chu, G.D.; Liang, R.; Wan, C.M.; Yang, J.; Li, J.; Wang, R.N.; Du, L.N.; Lin, R.X. Ultrasonic-assisted extraction of flavonoids from Juglans mandshurica Maxim.: Artificial intelligence-based optimization, kinetics estimation, and antioxidant potential. Molecules 2022, 27, 4837. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lu, M.F.; Liao, H.B.; Li, Y.X.; Han, W.; Yuan, K. Content determination of the flavonoids in the different parts and different species of Abelmoschus esculentus L. by reversed phase-high performance liquid chromatograph and colorimetric method. Pharmacogn. Mag. 2014, 10, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xue, J.; Zhang, S.; Zheng, S.; Xue, Y.; Xu, D.; Zhang, X. Composition of peony petal fatty acids and flavonoids and their effect on Caenorhabditis elegans lifespan. Plant Physiol. Biochem. 2020, 155, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sdiri, M.; Li, X.M.; Du, W.W.; El-Bok, S.; Xie, Y.Z.; Ben-Attia, M.; Yang, B.B. Anticancer activity of Cynomorium coccineum. Cancers 2018, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Ning, H.; He, G.; Huang, Y.; Wu, Z.; Jin, L.; Jiang, X. Rottlerin inhibits cell growth, induces apoptosis and cell cycle arrest, and inhibits cell invasion in human hepatocellular carcinoma. Mol. Med. Rep. 2018, 17, 459–464. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetics Models | Equations | Dynamic Parameters |
|---|---|---|
| Intra-particle diffusion | ki = 0.4796 mg/(g·h1/2) C = 5.729 mg/g R2 = 0.6140 | |
| Pseudo-first-order | k1 = 0.4989 h−1 Qe = 0.07 mg/g R2 = 0.6239 | |
| Peudo-second-order | k2 = 1.2365 g/(mg·h) Qe = 7.04 mg/g R2 = 0.9999 |
| Models | T (°C) | Equations | Parameters | ||
|---|---|---|---|---|---|
| KL (mL/mg) | Qm (mg/g) | R2 | |||
| Langmuir | 25 | 14.69 | 12.61 | 0.9973 | |
| 35 | 7.28 | 12.38 | 0.9965 | ||
| 45 | 5.25 | 11.75 | 0.9960 | ||
| KF [(mg/g)(mL/mg)1/n] | 1/n | R2 | |||
| Freundlich | 25 | 13.34 | 0.2916 | 0.8417 | |
| 35 | 11.83 | 0.3525 | 0.8962 | ||
| 45 | 10.50 | 0.3819 | 0.9014 | ||
| KT (mL/mg) | BT (J/mol) | R2 | |||
| Temkin | 25 | 201.64 | 2.382 | 0.8911 | |
| 35 | 74.70 | 2.622 | 0.9414 | ||
| 45 | 49.36 | 2.598 | 0.9487 | ||
| Group of Flavonoids | Structure Backbone | Examples | |
|---|---|---|---|
| chalcones |  |  |  |
| naringenin chalcone | cardamonin | ||
| dihydrochalcones |  |  |  |
| trilobatin | phloretin | ||
| flavanones |  |  |  |
| butin | eriodictyol | ||
| flavones |  |  |  |
| luteolin | apigenin | ||
| flavonols |  |  |  |
| kaempferol | quercitrin | ||
| anthocyanins |  |  |  |
| peonidin chloride | idaein chloride | ||
| isoflavonoids |  |  |  |
| puerarin | sophoricoside | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Chu, G.; Wu, J.; Zhang, G.; Du, L.; Lin, R. Enrichment and Evaluation of Antitumor Properties of Total Flavonoids from Juglans mandshurica Maxim. Molecules 2024, 29, 1976. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29091976
Yang S, Chu G, Wu J, Zhang G, Du L, Lin R. Enrichment and Evaluation of Antitumor Properties of Total Flavonoids from Juglans mandshurica Maxim. Molecules. 2024; 29(9):1976. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29091976
Chicago/Turabian StyleYang, Shuli, Guodong Chu, Jiacheng Wu, Guofeng Zhang, Linna Du, and Ruixin Lin. 2024. "Enrichment and Evaluation of Antitumor Properties of Total Flavonoids from Juglans mandshurica Maxim" Molecules 29, no. 9: 1976. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29091976