
A Metabolomics Study of the Effects of Eleutheroside B on Glucose and Lipid Metabolism in a Zebrafish Diabetes Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. EB Improves Disordered Glucose and Lipid Metabolism in Zebrafish
2.2. EB Increases the Expression of the GLUT4 Protein and Alleviates Diabetes Complicated with Liver Injury
2.3. Differential Metabolite Analysis
2.4. Metabolic Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Drugs and Materials
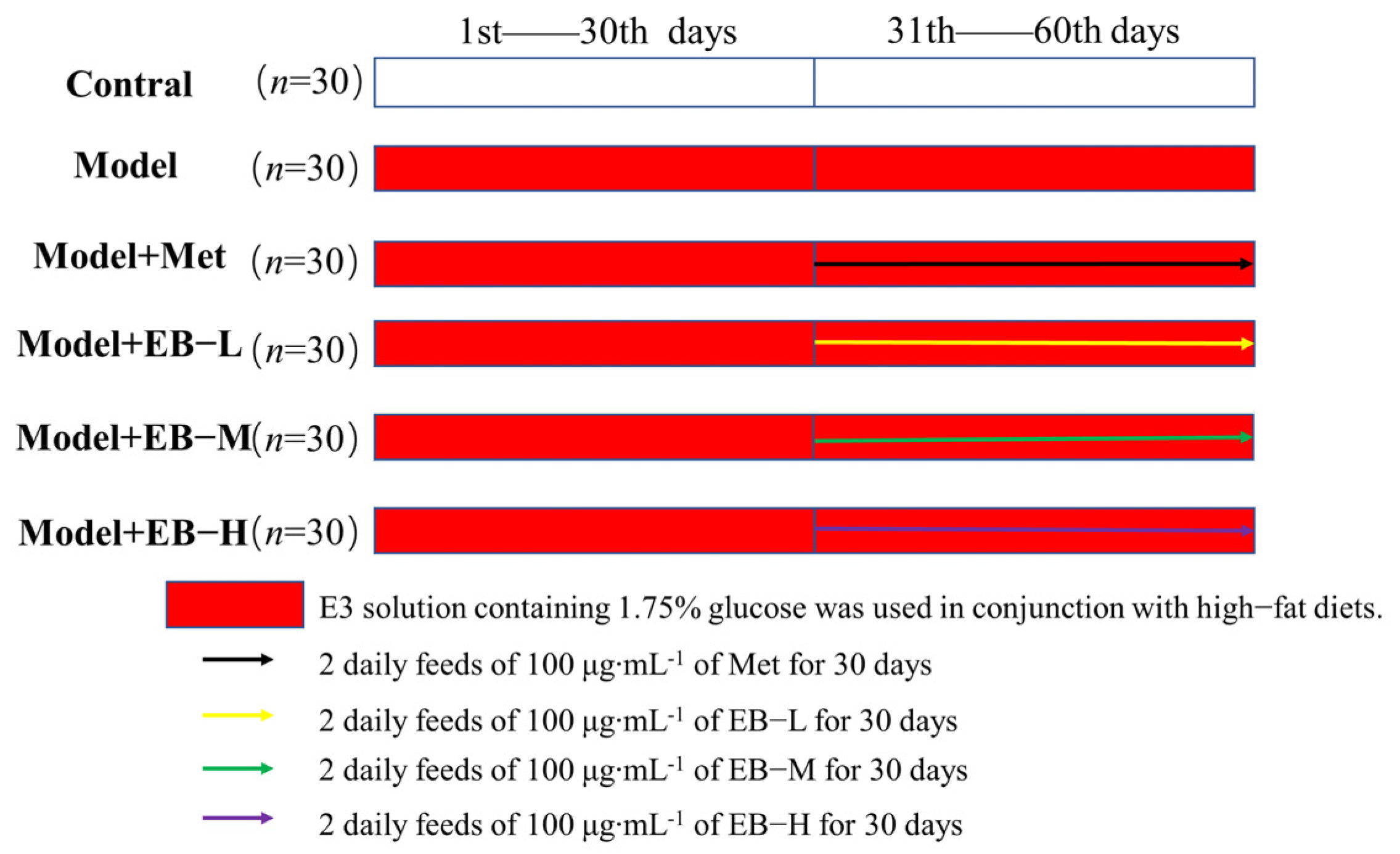
4.2. Animals and Experimental Design
4.3. Sample Collection
4.4. Measurement of Biochemical Indexes
4.5. Western Blot
4.6. Histopathological Examination
4.7. Metabolite Extraction
4.8. LC-MS Analysis
4.9. Data Anlaysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Satin, L.; Soleimanpour, S.; Walker, E. New Aspects of Diabetes Research and Therapeutic Development. Pharmacol. Rev. 2021, 73, 1001–1015. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9 edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, F.; Rorsman, P. Diabetes mellitus and the β cell: The last ten years. Cell 2012, 148, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, E.; Lim, S.; Lamptey, R.; Webb, D.; Davies, M. Type 2 diabetes. Lancet 2022, 400, 1803–1820. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Ma, R. Metabolomics in Diabetes and Diabetic Complications: Insights from Epidemiological Studies. Cells 2021, 10, 2832. [Google Scholar] [CrossRef] [PubMed]
- International Hypoglycaemia Study Group. Hypoglycaemia, cardiovascular disease, and mortality in diabetes: Epidemiology, pathogenesis, and management. Lancet Diabetes Endocrinol. 2019, 7, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Shulman, G. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef] [PubMed]
- Demir, S.; Nawroth, P.; Herzig, S.; Ekim Üstünel, B. Emerging Targets in Type 2 Diabetes and Diabetic Complications. Adv. Sci. 2021, 8, e2100275. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, L.; Wu, H.; Zhang, L.; Hu, X.; Li, C.; Liu, S. Noni (Morinda citrifolia L.) fruit phenolic extract supplementation ameliorates NAFLD by modulating insulin resistance, oxidative stress, inflammation, liver metabolism and gut microbiota. Food Res. Int. 2022, 160, 111732. [Google Scholar] [CrossRef]
- Gao, Q.; Li, G.; Zu, Y.; Xu, Y.; Wang, C.; Xiang, D.; He, W.; Shang, T.; Cheng, X.; Liu, D.; et al. Ginsenoside Rg1 alleviates ANIT-induced cholestatic liver injury by inhibiting hepatic inflammation and oxidative stress via SIRT1 activation. J. Ethnopharmacol. 2024, 319, 117089. [Google Scholar] [CrossRef]
- Zhou, N.; Zhao, Y.; Zhang, L.; Ning, Y. Protective effects of black onion polysaccharide on liver and kidney injury in T2DM rats through the synergistic impact of hypolipidemic and antioxidant abilities. Int. J. Biol. Macromol. 2022, 223, 378–390. [Google Scholar] [CrossRef]
- Saraswathi, V.; Kumar, N.; Gopal, T.; Bhatt, S.; Ai, W.; Ma, C.; Talmon, G.; Desouza, C. Lauric Acid versus Palmitic Acid: Effects on Adipose Tissue Inflammation, Insulin Resistance, and Non-Alcoholic Fatty Liver Disease in Obesity. Biology 2020, 9, 346. [Google Scholar] [CrossRef]
- Zoncu, R.; Efeyan, A.; Sabatini, D. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, Y.; Cao, X.; Peng, Y.; Huang, J.; Chen, L.; Pang, J.; Jiang, Z.; Qian, S.; Liu, Y.; et al. Targeting mTOR/YY1 signaling pathway by quercetin through CYP7A1-mediated cholesterol-to-bile acids conversion alleviated type 2 diabetes mellitus induced hepatic lipid accumulation. Phytomed. Int. J. Phytother. Phytopharm. 2023, 113, 154703. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sui, Y.; Wu, Q.; Xie, B.; Sun, Z. Attenuated mTOR Signaling and Enhanced Glucose Homeostasis by Dietary Supplementation with Lotus Seedpod Oligomeric Procyanidins in Streptozotocin (STZ)-Induced Diabetic Mice. J. Agric. Food Chem. 2017, 65, 3801–3810. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Balan, P.; Popovich, D. Review of Ginseng Anti-Diabetic Studies. Molecules 2019, 24, 4501. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Ata, A.; Anil Kumar, N.V.; Sharopov, F.; Ramírez-Alarcón, K.; Ruiz-Ortega, A.; Abdulmajid Ayatollahi, S.; Tsouh Fokou, P.; Kobarfard, F.; Amiruddin Zakaria, Z.; et al. Antidiabetic Potential of Medicinal Plants and Their Active Components. Biomolecules 2019, 9, 551. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Jamialahmadi, T.; Moallem, S.; Sahebkar, A. Boosting GLP-1 by Natural Products. Adv. Exp. Med. Biol. 2021, 1328, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Jia, A.; Zhang, Y.; Gao, H.; Zhang, Z.; Zhang, Y.; Wang, Z.; Zhang, J.; Deng, B.; Qiu, Z.; Fu, C. A review of Acanthopanax senticosus (Rupr and Maxim.) harms: From ethnopharmacological use to modern application. J. Ethnopharmacol. 2021, 268, 113586. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Q.; Yang, J.; Kan, M.; Jin, R.; Pu, T.; Yang, Y.; Xing, T.; Meng, X.; Zang, H. Eleutheroside B-loaded poly (lactic-co-glycolic acid) nanoparticles protect against renal fibrosis via Smad3-dependent mechanism. Phytother. Res. PTR 2021, 35, 6401–6416. [Google Scholar] [CrossRef]
- Che, D.; Zhao, B.; Fan, Y.; Han, R.; Zhang, C.; Qin, G.; Adams, S.; Jiang, H. Eleutheroside B increase tight junction proteins and anti-inflammatory cytokines expression in intestinal porcine jejunum epithelial cells (IPEC-J2). J. Anim. Physiol. Anim. Nutr. 2019, 103, 1174–1184. [Google Scholar] [CrossRef]
- Wang, F.; Yuan, C.; Liu, B.; Yang, Y.; Wu, H. Syringin exerts anti-breast cancer effects through PI3K-AKT and EGFR-RAS-RAF pathways. J. Transl. Med. 2022, 20, 310. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, Y.; Liu, G. Syringin may exert sleep-potentiating effects through the NOS/NO pathway. Fundam. Clin. Pharmacol. 2015, 29, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Us, M.; Zin, T.; Sundaram C., S.; Iqbal, M. Effect of syringin (eleutheroside B) on the physiological and hematological parameters in STZ induced Type II diabetic Wistar rats. Pak. J. Pharm. Sci. 2020, 33, 2601–2606. [Google Scholar] [PubMed]
- Yao, J.; Li, Y.; Jin, Y.; Chen, Y.; Tian, L.; He, W. Synergistic cardioptotection by tilianin and syringin in diabetic cardiomyopathy involves interaction of TLR4/NF-κB/NLRP3 and PGC1a/SIRT3 pathways. Int. Immunopharmacol. 2021, 96, 107728. [Google Scholar] [CrossRef] [PubMed]
- Vigna, L.; Tirelli, A.; Gaggini, M.; Di Piazza, S.; Tomaino, L.; Turolo, S.; Moroncini, G.; Chatzianagnostou, K.; Bamonti, F.; Vassalle, C. Insulin resistance and cardiometabolic indexes: Comparison of concordance in working-age subjects with overweight and obesity. Endocrine 2022, 77, 231–241. [Google Scholar] [CrossRef]
- Zhai, Z.; Yang, Y.; Lin, G.; Lin, W.; Wu, J.; Liu, X.; Zhang, S.; Zhou, Q.; Liu, H.; Hao, G. The hypertension and hyperlipidemia status among type 2 diabetic patients in the community and influencing factors analysis of glycemic control. Diabetol. Metab. Syndr. 2023, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Herman, R.; Kravos, N.; Jensterle, M.; Janež, A.; Dolžan, V. Metformin and Insulin Resistance: A Review of the Underlying Mechanisms behind Changes in GLUT4-Mediated Glucose Transport. Int. J. Mol. Sci. 2022, 23, 1264. [Google Scholar] [CrossRef] [PubMed]
- Kautzky-Willer, A.; Harreiter, J.; Pacini, G. Sex and Gender Differences in Risk, Pathophysiology and Complications of Type 2 Diabetes Mellitus. Endocr. Rev. 2016, 37, 278–316. [Google Scholar] [CrossRef]
- Perry, R.; Camporez, J.; Kursawe, R.; Titchenell, P.; Zhang, D.; Perry, C.; Jurczak, M.; Abudukadier, A.; Han, M.; Zhang, X.; et al. Hepatic acetyl CoA links adipose tissue inflammation to hepatic insulin resistance and type 2 diabetes. Cell 2015, 160, 745–758. [Google Scholar] [CrossRef]
- Wu, H.; Lu, Z.; Chen, R.; Cai, Q.; Wang, M.; Zhang, L.; Zhu, Z. Factors associated with gastrointestinal side effects after liraglutide treatment for type 2 diabetes. Front. Endocrinol. 2023, 14, 1098032. [Google Scholar] [CrossRef] [PubMed]
- Komajda, M.; McMurray, J.; Beck-Nielsen, H.; Gomis, R.; Hanefeld, M.; Pocock, S.; Curtis, P.; Jones, N.; Home, P. Heart failure events with rosiglitazone in type 2 diabetes: Data from the RECORD clinical trial. Eur. Heart J. 2010, 31, 824–831. [Google Scholar] [CrossRef]
- Okayasu, S.; Kitaichi, K.; Hori, A.; Suwa, T.; Horikawa, Y.; Yamamoto, M.; Takeda, J.; Itoh, Y. The evaluation of risk factors associated with adverse drug reactions by metformin in type 2 diabetes mellitus. Biol. Pharm. Bull. 2012, 35, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Yuxin, H.; Cuiping, J.; Wen, T.; Jieyuzhen, Q.; Xiaoming, T.; Qin, G.; Haidong, W.; Jiao, S.; Zhijun, B. Comparison of gastrointestinal adverse events with different doses of metformin in the treatment of elderly people with type 2 diabetes. J. Clin. Pharm. Ther. 2020, 45, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chen, Q.; Liu, Y.; Jin, L.; Peng, R. Research Progress on the Construction and Application of a Diabetic Zebrafish Model. Int. J. Mol. Sci. 2023, 24, 5195. [Google Scholar] [CrossRef]
- Jones, J. Hepatic glucose and lipid metabolism. Diabetologia 2016, 59, 1098–1103. [Google Scholar] [CrossRef]
- Wang, L.; Yan, N.; Zhang, M.; Pan, R.; Dang, Y.; Niu, Y. The association between blood glucose levels and lipids or lipid ratios in type 2 diabetes patients: A cross-sectional study. Front. Endocrinol. 2022, 13, 969080. [Google Scholar] [CrossRef]
- Johnson, A.; Olefsky, J. The origins and drivers of insulin resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Różańska, D.; Regulska-Ilow, B. The significance of anthocyanins in the prevention and treatment of type 2 diabetes. Adv. Clin. Exp. Med. 2018, 27, 135–142. [Google Scholar] [CrossRef]
- Alsahli, M.A.; Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Anwar, S.; Almutary, A.G.; Alrumaihi, F.; Rahmani, A.H. 6-Gingerol, a Major Ingredient of Ginger Attenuates -Induced Liver Injury in Rats through the Modulation of Oxidative Stress and Anti-Inflammatory Activity. Mediat. Inflamm. 2021, 2021, 6661937. [Google Scholar] [CrossRef]
- Newgard, C. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Z.; Wang, D.; Jiang, Y.; Liu, Y. Targeting mTOR Signaling in Type 2 Diabetes Mellitus and Diabetes Complications. Curr. Drug Targets 2022, 23, 692–710. [Google Scholar] [CrossRef]
- Ramasubbu, K.; Devi Rajeswari, V. Impairment of insulin signaling pathway PI3K/Akt/mTOR and insulin resistance induced AGEs on diabetes mellitus and neurodegenerative diseases: A perspective review. Mol. Cell. Biochem. 2023, 478, 1307–1324. [Google Scholar] [CrossRef] [PubMed]
- Gökcen, B.; Şanlier, N. Coffee consumption and disease correlations. Crit. Rev. Food Sci. Nutr. 2019, 59, 336–348. [Google Scholar] [CrossRef]
- Eny, K.; Orchard, T.; Miller, R.; Maynard, J.; Grant, D.; Costacou, T.; Cleary, P.; Braffett, B.; Paterson, A. Caffeine Consumption Contributes to Skin Intrinsic Fluorescence in Type 1 Diabetes. Diabetes Technol. Ther. 2015, 17, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Guo, J.; Chen, H.; Zhang, J.; Zhang, X.; Jiang, X.; Wang, F.; Xu, Z.; Huang, X.; Sha, J.; et al. Maternal caffeine exposure impairs insulin secretion by pancreatic β-cells and increases the risk of type II diabetes mellitus in offspring. Cell Biol. Int. 2014, 38, 1183–1193. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, X.; Chen, Q.; Chi, W.; Qiu, Z.; Qiu, Y. A Metabolomics Study of the Effects of Eleutheroside B on Glucose and Lipid Metabolism in a Zebrafish Diabetes Model. Molecules 2024, 29, 1545. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29071545
Dong X, Chen Q, Chi W, Qiu Z, Qiu Y. A Metabolomics Study of the Effects of Eleutheroside B on Glucose and Lipid Metabolism in a Zebrafish Diabetes Model. Molecules. 2024; 29(7):1545. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29071545
Chicago/Turabian StyleDong, Xuelian, Qiang Chen, Wenyan Chi, Zhidong Qiu, and Ye Qiu. 2024. "A Metabolomics Study of the Effects of Eleutheroside B on Glucose and Lipid Metabolism in a Zebrafish Diabetes Model" Molecules 29, no. 7: 1545. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules29071545